Zootecnica generale appunti
Zootecnica generale appunti
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Le informazioni di medicina e salute contenute nel sito sono di natura generale ed a scopo puramente divulgativo e per questo motivo non possono sostituire in alcun caso il consiglio di un medico (ovvero un soggetto abilitato legalmente alla professione).
Zootecnica generale appunti
APPUNTI DEL CORSO DI ZOOTECNICA GENERALE
1. I SISTEMI ZOOTECNICI
Le modalità con cui si esercita la pratica dell’all evamento animale sono diverse, ma riconducibili ad un idea di sistema zootecnico.
Per sistema zootecnico s’intende l’organizzazione delle entità che compong ono l’ allevamento e che ne rappresentano le variabili di stato (ambiente fisico, animali, produzioni, mezzi tecnici, lavoro, capitali), e delle loro relazioni che rappresentano le funzioni (transiti di energia, materiali e informazioni).
E’ difficile fare una classificazione dei sistemi zootecnici, perchè le aree di confine fra i sistemi sono aleatorie ed i criteri in base ai quali formuliamo la classificazione possono essere arbitrari.
Possiamo effettuare la classificazione dei sistemi zootecnici secondo quattro criteri guida:
1) Produttività e legame con la base territoriale agri cola dal quale dipende il rifornimento degli
alimenti zootecnici impiegati nel ciclo produttivo: l’ allevamento animale può dipendere in toto, parzialmente oppure essere slegato dalla base territoriale di riferimento per la produzione degli alimenti. In funzione della produttività possiamo distinguere colture o superfici foraggiere ad alto, medio o basso livello produttivo. Se tutti gli alimenti impiegati sono acquisiti dall’esterno, gli allevamenti prendono il nome di imprese senza terra.
La produttività delle colture foraggiere zootecnich e è legata alla fertilità del suolo, alle condizioni ambientali, agli interventi tecnici quali irrigazione, concimazione, lavorazioni, difesa da malerbe e parassiti. Un indicatore indiretto di produttività è il carico animale mantenibile che rappresenta il numero di capi ( espressi in unità convenzionali o in kg di peso corporeo mantenuto) per unità di superficie (normalmente l’ettaro) in condizioni ordinarie (ovvero in un’annata media). Tale carico deve essere calcolato solo sulle risorse foraggiere aziendali, escludendo gli alimenti acquistati dall’esterno.
Gli animali occupano una base territoriale virtuale in quanto gli alimenti che consumano sono, in ogni caso, derivati dalla coltivazione di una porzione di terreno appartenente alla stessa azienda nella quale essi sono allevati oppure in aziende lontane (a volte dislocate in altri continenti). La quantità di a limenti provenienti dalla azienda in cui risiede l’allevamento rapportata al consumo complessivo alimentare annuale (espresso in termini di energia o di s.s.) costituisce l’indice di autoapprovvigionamento.
- specializzazione aziendale e livello produttivo degli animali: dobbiamo distinguere le aziende specializzate, dove viene allevata una sola specie normalmente con un solo indirizzo produttivo (esbovini per produzione di latte o di carne); da quelle miste nelle quali sono presenti più specie; il livello produttivo, riferito alla media della popolazione per quel carattere, viene espresso in percentuale, ed individua la posizione nella quale si colloca l’azienda rispetto alla produttività della popolazione d i riferimento.
- grado di organizzazione interna: possiamo distinguere le imprese zootecniche in base al livello delle tecnologie impiegate e possiamo distinguere in allevamenti dotati di elevato livello tecnologico (uso dell’ I.A, diagnosi di gravidanza, computerizzazione di alcune operazioni, elevato livello di meccanizzazione), da quelli a medio o a basso livello tecnologico nei quali sono adottate mediamente oppure episodicamente tecniche avanzate di produzione. Questo criterio si riferisce anche alle modalità di rilevazione e di elaborazione dei dati aziendali, ciò è importante in quanto l’azienda zootecnica è quella che, nel settore agricolo, produce la maggior mole di dati e la capacità di raccoglierli ed elaborarli in tempo reale è di importanza fondamentale affinché l’imprenditor possa prendere tempestivamente le decisioni tecniche e gestionali sulla conduzione dell’impresa.
- aspetti socio-economici: Possiamo classificare gli allevamenti in funzione del contesto sociale ed economico in cui essi sono collocati. Il tipo d’impresa ed i suoi collegamenti con il mercato nazionale o internazionale, sia per la vendita dei prodotti che per l’approvvigionamento di mezzi tecnici e capitali, sono rilevanti ai fini della sua collocazione nel contesto produttivo. Es. nei paesi in via di sviluppo nei alcune forme di allevamento sono praticate con la finalità di tesaurizzazione del capitale bestiame.
La competizione fra le imprese in regime di concorrenza porta ad un aumento d’intensità produttiva al fine di ridurre il costo unitario di produzione. Però il costo unitario può anche essere ridotto con u n abbassamento del grado d’impiego dei fattori produttivi: tale concetto è uno dei perni della politica dell’Unione Europea per gli anni 2000-2006 (contenuta nella cosiddetta Agenda 2000) ed è indicata con il termine di estensivizzazione. In particolare l’estensivizzazione proposta dalla PAC tende a ridurre il carico unitario (espresso in Unità Bovin e Adulte UBA, una misura standardizzata della consistenza animale) e ad utilizzare la risorsa foraggera disponibile in modo sub-ottimale.
ICL: il grado di intensità di una impresa è il rapporto fra il costo del lavoro e il fatturato definito come indice costo del lavoro
Sulla base dei criteri sopraddetti possiamo classificare gli allevamenti in quattro tipi di sistema produttivo a seconda dell’intensità con cui sono im piegati i fattori della produzione (terra, lavoro, capitale, organizzazione).
- Intensivo. Questo sistema d’allevamento è di norma specializzato, con un unico indirizzo produttivo principale (eventualmente uno secondario come nel caso in cui si produca latte, ma si vendano anche gli animali per il ristallo = ingrassamento o per la macellazione), ad elevato livello produttivo, con alta tecnologia e ben calato in un ambiente imprenditoriale maturo e ampiamente collegato. L’ICL è compreso fra il 20 e il 30%; l’alimentazione rappresentata la maggior parte dei costi. Rientrano in questa categoria le aziende d’allevamento senza terra di suini, avicoli e conigli nelle quali l’intero ammontare degli alimenti è acquisita dal mercato. Gli animali sono in genere mantenuti in stabulazione permanente.
- Semintensivo. I fattori della produzione sono impiegati ad un livello inferiore rispetto al precedente. Gli animali sono mantenuti in stabulazione semipermanente e l’ICL è compreso fra il 30 ed il 40%. Es. sono gli allevamenti di bovine da latte in cui è praticato anche il pascolamento, allevamenti avicoli a terra, suini plein air, i sistemi semistabulati di allevamento degli ovini e dei caprini da latte (parziale ricorso al pascolamento).
- Semiestensivo. E’ normalmente praticato con ampio ricorso al pascolamento. Si tratta solitamente di allevamenti misti, con livello produttivo medio-basso e con un ICL compreso fra il 40 ed il 50%. Rientrano in questa categoria gli allevamenti ovini da latte e da carne di buon livello produttivo, gli allevamenti bovini all’alpeggio.
- Estensivo.Tutti i fattori della produzione sono utilizzati al livello minimo. Il ricovero degli animali è inesistente oppure saltuario. La risorsa alimentare prevalente o esclusiva è il pascolo, l’indirizzo produttivo è di solito misto, i carichi unitari sono piuttosto bassi. é il tipico sistema d’allevamento degli ovini da lana e da carne, dei bovini rustici, dei caprini e dei suini allo stato brado.E’ diffuso in vaste aree dei Paesi in via di sviluppo, ma è frequente anche in zone dei paesi sviluppati non diversamente valorizzabili. Da qualche tempo si indica questo tipo di allevamento come marginaleper indicare una situazione territoriale caratterizzata da forti limiti sotto il profilo della morfologia, della fertilità dei suoli, del clima e delle infrastrutture, anche gli allevamenti che vi si praticano sono detti i marginali.

2. RICHIAMI DI FISIOLOGIA DELLA DIGESTIONE
MICROBICA E CENNNI SULL’INGESTIONE E SUL
COMPORTAMENTO ALIMENTARE
2.1. PREMESSA
In tutte le specie zootecniche la digestione microbica è più o meno importante nell' utilizzazione nutritiva degli alimenti.
Nel canale digerente di tutti gli animali, infatti, è sempre presente una certa attività microbica, che è più importante nelle specie erbivore, meno in quelle onnivore e marginale in quelle carnivore.
L’attività dei microbi nelle specie erbivore è indi spensabile. I microorganismi alberganti nel
digerente infatti:
- consentono l’attacco e la degradazione della parete della cellula vegetale (fibra), la che risulterebbe non digeribile in quanto gli animali non hanno gli enzimi capaci di attaccare i carboidrati strutturali (emicellulose, cellulosa);
- producono acidi grassi volatili (AGV) utilizzati dall’animale nel proprio metabolismo;
- sintetizzano proteine ad elevato valore biologico;
- sintetizzano vitamine del gruppo B.
L’attività microbica può essere:
- prepeptica: è avviene anteriormente alla digestione gastrica (ruminanti e pseudoruminanti);
- postpeptica: è avviene posteriormente alla digestione gastrica (equidi);
3) interpeptica: è intermedia alla digestione gastrica (ciecotrofi = conigli).
PREPETICA: la fermentazione degli alimenti avviene in un sacco prestomacale (rumine, per
i bovini, o pseudorumine, per i camelidi) ed è accompagnata da una masticazione secondaria rispetto a quella ingestiva. Il 70% delle sostanze nutritive sono utilizzate in questa sede e la digestione gastrica riguarda prevalentemente le proteine. Questa soluzione consente la migliore utilizzazione della fibra e dell’azoto non proteico, ma non è ottimale l’utilizzazione dei carboidrati di riserva (CNS = carboidrati non strutturali, quali zuccheri semplici e amido).
POSTPEPTICA: la digestione della fibra avviene nel grosso colon, dopo che gli amidi e le proteine sono state soggette alla digestione gastrica e intestinale. Questa soluzione consente la migliore
utilizzazione dei CNS, ma una minore degradazione della fibra rispetto a quella precedente. Infatti, in presenza di alimenti poveri (ricchi in fibra e poveri in CNS), gli equidi devono ingerire una quantità superiore rispetto ai ruminanti per trarne la stessa quantità di energia.
INTERPEPTICA: la digestione microbica avviene nel grosso colon con la produzione di ciecotrofo, che è una pallottola mucillaginosa, in stato di fermentazione, di odore pungente; queste digesta sono emesse dall’ano del coniglio (o lepre) , reingerite e ridigerite nello stomaco e nell’intestino tenue . Alla fine di tale processo avviene l’espulsione delle feci vere (praticamente inodori) per via anale. La ciecotrofia è il sistema digestivo degli erbivori più efficiente. I componenti degli alimenti digeribili per via enzimatica dall’animale sono infatti digeriti prima che inizi la fermentazione microbica che attacca la fibra e i composti non digeriti con la produzione di AGV; la seconda digestione gastro-intestinale serve per digerire i prodotti non volatili ottenuti dalla fermentazione microbica.
Il più importante è il primo perchè tipico dei ruminanti, che comprendono la parte più rilevante delle specie allevate: i bovini, i bufalini, i caprini e gli ovini.
2.2. RITENZIONE DEGLI ALIMENTI NEL RUMINE- RUOLO DELLA MASTICAZIONE
2.2.1 Il contenuto del rumine
Nel rumine-reticolo è presente in continuazione una massa alimentare fibrosa in corso di fermentazione che rappresenta all’incirca i ¾ del contenuto diges tivo totale e dall’8 al 17% del peso corporeo (PC) totale dell’animale, in funzione del tipo della razione somministrata. L’acqua rappresenta l’80-90% de l contenuto fresco del rumine e le pareti cellulari vegetali(CS = carboidrati strutturali determinati analiticamente dalla fibra al digerente neutro =NDF) più dell’80% del contenuto secco.
Il rumine è approvvigionato durante 5-8 ore al giorno con gli alimenti che vengono ingeriti in una dozzina di pasti. Gli animali al pascolo effettuano due grandi pasti, uno all’alba e l’altro alla sera ; anche gli animali in stabulazione, sia fissa che libera.
Il contenuto ruminale si accresce durante i pasti ed arriva al massimo durante il grande pasto della sera. L’acqua è apportata dagli alimenti, dall’abbeverata e dalla saliva che è secreta sia continuamente dalle ghiandole parotidi, che nel corso dell’ingestione e della ruminazione (ghiandole sottolinguali): circa 150 l/ gg nel bovino adulto e 10 l/gg negli ovini adulti. La saliva è leggermente alcalina (pH 8,2) e ricca in sostanze tampone (bicarbonati e fosfati) che contribuiscono a mantenere la reazione del mezzo ruminale ad un pH tra 6,2 a 6,5. Il contenuto ruminale è molto condizionato dalla temperatura corporea
e dell’anaerobiosi: infatti, l’efficienza delle fer mentazioni è massima in totale assenza di ossigeno, con temperatura costante, con un pH subacido e con un sistema dotato in continuo sia dell’alimentazione che della rimozione dei prodotti della fermentazione.
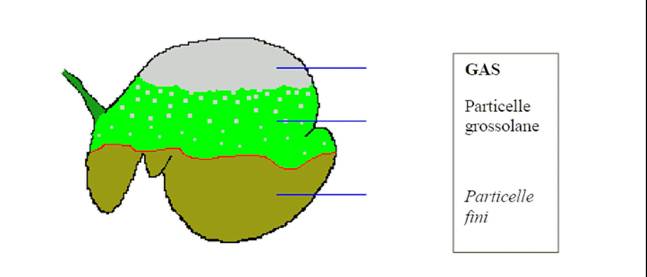
Figura 1.1 . Rappresentazione schematica della stratificazione dei componenti del contenuto del rumine.
Il contenuto ruminale è rimescolato permanentemente attraverso circuiti obbligatori attivati da circa 2500 contrazioni giornaliere che percorrono la parete ruminale, i pilastri e le pliche di separazione fra i diversi sacchi. Le contrazioni principali, primarie, partono dal reticolo (durata 4 secondi) al ritmo di 1,5 circa al minuto durante i pasti e di uno al minuto nel restante tempo. Si dirigono verso la parte posteriore dell’organo e sono spesso seguite da contrazioni secondarie le quali partono dal fondo del sacco ventrale e si dirigono in senso opposto verso la parte anteriore dello stesso.
Il flusso di alimenti e di liquidi implica lo svuotamento del rumine; gli alimenti scompaiono per due vie:
- La distruzione quasi totale dei tessuti e delle pareti non lignificate da parte della microflora albergante accompagnata dall’evacuazione dei prodotti terminali della loro fermentazione (assorbimento degli AGV ed eruttazione della CO2 e del metano);
- L’evacuazione verso valle attraverso l’orifizio reticolo-omasale (ORO) delle particelle fibrose non degradate, costituite da tessuti lignificati, , accompagnate dalla massa batterica in fermentazione (gli alimenti che realmente digerisce il ruminante sono quelli che lasciano il rumine indigeriti, il
cosiddetto escape ruminale , e dai corpi dei batteri e dei protozoi che albergano nel rumine e che hanno colonizzato le particelle alimentari o che nuotano liberamente nel fluido ruminale).
La dimensione di questo orifizio è piccola, ma varia in maniera ciclica; esso si dilata improvvisamente alla fine della contrazione del reticolo e forma un’apertura ellittica lunga 4 cm e larga 1 cm al massimo nei grossi bovini. E durante questo periodo di apertura dell’orifizio, che si ripete per circa 1600 volte al giorno, che si effettua l’evacuazione del contenuto ruminale. Questo particolare dell’orifizio reticolo-omasale, e i meccanismi della frantumazione degli alimenti per mezzo della masticazione mericica, fa passare solo le particelle < 1-2 mm negli ovini e 2-4 mm nei bovini. In generale le particelle presenti nell’omaso sono inferiori al millimetro e non subiranno una ulteriore riduzione di dimensione nel corso della successiva digestione.
2.2.2 Circuito di frantumazione degli alimenti: ruolo della ruminazione
I ruminanti masticano gli alimenti nel corso dell’ingestione (masticazione ingestiva) con una velocità che è 2 volte più elevata nei piccoli ruminanti: 125-150 movimenti mandibolari per minuto negli ovini e 70-80 movimenti nei bovini
Il tempo di masticazione per kg di sostanza secca è basso per gli alimenti concentrati, in quanto essi sono stati macinati più o meno finemente nel corso della preparazione. Esso è tanto più lunga per i foraggi più sono più fibrosi: circa 30 minuti per un buon fieno, oltre 60 minuti per una paglia, 20 minuti per l’insilato di mais contro 5-10 minuti per i concentrati, nel caso della vacca da latte.
La masticazione ingestiva riduce gli alimenti in un insieme di particelle di dimensione e di forma estremamente variabili, da un minimo di 1 mm ad un massimo di 4-5 cm. Le piccole particelle (<1 mm) rappresentano circa il 15% della sostanza secca dei foraggi coriacei e intorno alla metà nei foraggi teneri.
I frammenti alimentari originatisi da questa masticazione, vengono ingeriti con un fiotto di saliva e costituiscono il bolo alimentare. Essi sono energicamente sospinti verso la porzione caudale del rumine dalle contrazioni del reticolo quale sia la loro dimensione, e sono costretti ad immergersi nella massa fibrosa presente nell’organo. Tale immersione è tanto più rapida quanto più piccoli, più densi e più idrofori sono i frammenti; essa è più rapida per i concentrati e più lenta per i foraggi secchi. Il contenuto ruminale alimentare con razioni costituite da foraggi secchi, è nettamente stratificato: sotto una cupola di gas, si ha uno strato di particelle di foraggi più grandi e leggeri; nel sacco ventrale si ha invece un contenuto più fluido con particelle più piccole e più dense.
Poco dopo la fine di un pasto, di solito da 5 a 15 minuti, per l’animale inizia un periodo di ruminazione che è la successione di cicli la cui durata è di circa 1 minuto. Ciascun ciclo inizia con una contrazione supplementare del reticolo, che precede la sua contrazione primaria di qualche secondo. Un bolo (50-80
gr nell’ovino e 600-1000 gr nel bovino) derivante del contenuto del reticolo o del sacco craniale del rumine, è aspirato dall’esofago e rimonta fino alla bocca (bolo mericico). Se il bolo è troppo grosso, la parte eccedente è reingerita ed il rimanente subisce una intensa masticazione (masticazione mericica) che è effettuata con 40-60 movimenti di mandibola al minuto nei bovini e 80-100 negli ovini. In tal modo, le grosse particelle sono ridotte di dimensione e trasformate in piccole particelle (<1mm). Con razioni normali a base di foraggi la ruminazione occupa più tempo dell’ingestione e si effettua in un numero maggiore di periodi; la quantità di s. s. co sì masticata al giorno nel corso della ruminazione è 2 o 3 volte superiore a quella ingerita.
La masticazione mericica è importante nella frantumazione delle particelle alimentari e nel loro tempo di ritenzione ruminale. Con la riduzione della taglia e l’aumento della densità e la perdita delle par ti degradabili le particelle hanno una probabilità sem pre maggiore di sfuggire alla massa fibrosa, di immergersi verso il sacco ventrale e di guadagnare la parte inferiore di quello craniale da cui sono sospinte verso orifizio reticolo omasale.
Figura 1.2 . Circuito delle particelle alimentari nel rumine.
La masticazione mericica permette lo svuotamento del rumine che è necessario per un suo successivo riempimento. Le fermentazioni ruminali, rendendo meno resistenti le pareti cellulari e privando le particelle della frazione degradabile, accrescono la loro efficacia.
2. 2.3 Tempi di masticazione, tempi di ritenzione, ingombro e ingeribilità.
I tessuti più resistenti alla masticazione, sia durante la ruminazione che ingestione, sono quelli lignificati: lo sclerenchima ed i vasi legnosi.
Il risultato è che per kg di sostanza secca ingerita si ha un aumento simultaneo:
- del tempo d’ingestione;
- del tempo di ruminazione, accentuato dal fatto che la popolazione microbica, che è alimentata meno bene per effetto della diminuzione del contenuto cellulare costituito dal CS e proteine, degrada più lentamente la parete cellulare;
- del tempo di ritenzione medio delle particelle nel rumine in quanto dipende dal tempo di ruminazione;
- dell’effetto d’ingombro esercitato dalla razione in quanto questo è prodotto dalla proporzione delle particelle non degradabili per il loro tempo di ritenzione, che aumentano entrambi;
- del peso del contenuto ruminale espresso in tal q uale o in sostanza secca.
Il tempo dedicato alla ruminazione non è mai superiore alle 11 ore al giorno; quando è vicino a tale valore può ridursi il tempo di ingestione in quanto il tempo totale di masticazione(ingestione + ruminazione) non può eccedere le 18 ore al giorno a causa dell’affaticamento dei muscoli masticatori e delle altre esigenze dell’animale.
Il tempo di ritenzione ruminale (MRT) esprime (in ore) la permanenza media di una determinata classe di particelle, o del liquido compresi i suoi soluti, nel rumine. Il ricambio delle parti solide e dei liquidi presenti nel rumine prende il nome di turnover ruminale. I MRT dei solidi è superiore a quello dei liquidi; esso è maggiore nei bovini rispetto ai piccoli ruminanti per i solidi, mentre quello dei liquidi è pressoché uguale fra le specie. Il MRT dei solidi rispetto ai liquidi è di circa 3 volte nei bovini e di 1,6-1,7 volte nei piccoli ruminanti. Il MRT dipende sia dalle dimensioni che dal peso specifico delle particelle alimentari. Al crescere della densità re lativa (g/ml) il MRT si riduce: ciò significa che l e particelle più pesanti hanno maggiore probabilità d i affondare nel rumine e di depositarsi sul fondo dal quale poi sono sospinte nel reticolo per passare oltre. Anche la dimensione è rilevante ai fini del MRT: si nota la riduzione del MRT con l’aumento della densità, ma con MRT sempre maggiore a parità di peso con particelle più grandi.
Il TMR nel caso di solidi è vacca> capre > pecore Il TMR nel caso di liquidi è capre >pecore> vacca
2.3 LE FERMENTAZIONI MICROBICHE
2.3.1 La micropopolazione ruminale.
Nel rumine è presente una imponente massa di microorganismi appartenenti ai gruppi dei batteri,
dei protozoi e dei funghi; per lama è anaerobio stretto, ma sono anche presenti delle specie e dei ceppi aerobi facoltativi che hanno il compito di assorbire l’ossigeno che penetra nelrumine con gli alimenti e quello che filtra dalla parete dell’organo.
I microorganismi, di solito hanno flagelli o ciglia mobili per nuotare nel liquido ruminale, colonizzano i frammenti di alimento che si trovano immersi nella massa in fermentazione.
La concentrazione batterica e quella protozoaria si riducono al crescere della dimensione del microorganismo.
All’ interno della popolazione microbica l’interazi one più frequente è la sequenzialità, dove un prodotto della fermentazione di una specie batterica è indispensabile per la sopravvivenza di una o più specie.
I batteri sono il gruppo più numeroso e preponderante della micropopolazione ruminale. Il loro compito è di colonizzare le particelle di piante ingerite dall’animale, attaccarle con esoenzimi, solubilizzare i CS e fermentarli fino ad AGV, attaccare le proteine alimentari e l’NPN per sintetizzare le proteine del loro corpo, sintetizzare le vitamine del gruppo B.
I protozoi non hanno un ruolo ben definito nel quadro delle fermentazioni ruminali. Predando i batteriaccrescono il valore biologico delle proteine che dal rumine passano alla digestione gastrica; il VB delle proteine dei protozoi è superiore a quello delle proteine del batteri; possono inglobare sia batteri in fase di fermentazione attiva, ma anche frazioni del contenuto ruminale, favorendo la fermentazione.
I funghi anaerobi presenti nel rumine possono essere sia parassiti dei protozoi ciliati che saprofiti deitessuti vegetali. Questi ultimi giocano un ruolo importante nell’aumentare le disponibilità dei conten uti intracellulari delle cellule vegetali per azione meccanica o enzimatica, anche solubilizzando parte della lignina (che non sarà digerita comunque). I funghi hanno un ruolo sinergico con i batteri nella produzione del metano.
Figura 1.4 . Schema della popolazione microbica del rumine.
Rappresentazione della popolazione batterica nel rumine dei bovini. I batteri si trovano nel fluido
ruminale, sulle particelle alimentari, e attaccate all’epitelio ruminale, dal quale le cellule distali cadono nel fluido. Sono mostrati i batteri aderenti al tessuto in relazione all’urea e all’ossigeno ch e si diffondono attraverso la parete dell’organo, e verso le cellule epiteliali distali, le quali comprendono il loro substrato nutritivo.
2.3.2. Le fermentazioni
I batteri del rumine colonizzano i substrati costituiti dagli alimenti e si accrescono. L’accrescimento della popolazione necessita energia, ottenuta dalla fermentazione dei carboidrati, e proteine che sono ottenute a partire dalle sostanze azotate. I lipidi, a causa della mancanza d’ossigeno, non possono essere impiegati dai batteri come fonte energetica. Tuttavia i grassi sono utilizzati costruire la membrana cellulare batterica e gli acidi grassi insaturi (maggioranza dei costituenti dei lipidi vegetali) sono saturati dai batteri in quanto sono per loro tossici.
Le fermentazioni sui carboidrati.
La degradazione ruminale dei carboidrati inizia con l’attacco da parte dei batteri dei principali CS (cellulose ed emicellulose) e CNS (amidi e zuccheri semplici) con enzimi extracellulari. Tale attacco porta alla formazione direttamente di glucosio (cellulosa, amidi) o di fruttosio (pectine, emicellulose, pentosani, fruttosani). Le cellule animali hanno una grande affinità per il glucosio in quanto present ano meccanismi per inglobarlo; le cellule della parete del rumine sono impermeabili al glucosio in modo da non sottrarlo al liquido ruminale e rendere possibile la successiva utilizzazione da parte dei batteri. Terminata la fase extracellulare, i monosaccaridi sono utilizzati dai batteri per il proprio metabolismo. Per primo i batteri demoliscono il fruttosio1-6 DI_P (6C) fino ad ac. piruvico (3C, si hanno 2 moli di piruvato per ogni mole di monosaccaride fermentato). L’ac. piruvico è pertanto l’intermediario della fermentazione.
Ora, se il ceppo (o i ceppi) batterico sta fermentando la cellulosa, o altri componenti della fibra (batteri cellulosolitici; fermentano bene ad un pH > 6,1), al termine producono ac. acetico, CO2 e ac. formico. L’acido formico fornisce il substrato di fermentazione per i metanobatteri, che sono importanti per il mantenimento della reazione del mezzo nei sistemi anaerobici, sono archeobatteri. Hanno il compito di ossidare l’idrogeno che si libera nel corso delle fermentazioni con il carbonio per produrre metano, con un meccanismo analogo alla fosforilazione ossidativa degli organismi aerobici, dove il metano è l’analogo dell’acqua dei sistemi aerobi. In definit iva, l’attacco ad una mole di esoso (6C) che deriva dalla degradazione dei carboidrati della parete cellulare (CS), porta alla produzione di 2 moli di acido acetico (2 x 2C = 4C), ad una mole di metano (1C) ed a una mole di anidride carbonica (1C). Il
guadagno energetico per ciascuna mole di esoso fermentato per questa via è di 4 ATP (1 per la glicolisi + 1 per la produzione di acetato, per mole di piruvato, essendoci 2 moli di piruvato per ogni mole di glucosio che entra nella catena). Il butirrato (4C) è prodotto dai batteri butirrici molto attivi ad un pH compreso fra il 6,3 e il 5,5 con una via iniziale simile a quella dei batteri acetici. Il prodotto finale però non è l’acetato in quanto questo ultimo è attivatoad ecetil-COA e, attraverso la β-condensazione (o la via alternativa del malonato), condensato in acido butirrico (2C + 2C = 4C). Anche in questo caso, per mole di esoso fermentato abbiamo la produzione di una mole di metano ed una mole di CO2. Il bilancio definitivo per mole di esoso è pertanto di una mole di butirrato (4C) + una mole di metano (1C) + una mole di CO2 (1C), con un guadagno energetico di 3 ATP (2 nella glicolisi ed 1 nella butirrogenesi).
I batteri che attaccano i substrati costituiti da CNS sono chiamati amilolitici, hanno un pH ottimale fra 6 e 5. La loro attività fermentativa porta alla pro duzione di acido propionico (3C) che, fra i 3 AGV, è quello meno forte (pKa 4,87 rispetto ad un valore di 4,82 per il butirrico e di 4,75 per l’acetico). Da una mole di esoso si ottengono 2moli di propionato: in questa reazione non si ha perdita di materia ne di energia, in quanto non si perde carbonio ne sotto forma di CO2 ne sotto forma di CH4. Tale via fermentativa fornisce un bilancio dell’ATP dovuto solamente alla glicolisi (2 ATP), ma ha il vantaggio di scaricare(ossidare) i sistemi di trasporto dell’idrogeno (NAD+, FAD+) che, in ambiente riducente come quello anaerobico, sono normalmente carichi (ridotti = NADH-H+; NADPH-H+).
Un quadro riassuntivo delle fermentazioni ruminali è riportato di seguito: si parte da 5 moli di carboidrati vegetali (30 C) per arrivare ad un rapporto molare fra i tre AGV tipico (67:22:11) nel caso di razioni medie: 6C sono trasformati in acetato; 2C in propionato; 1C in butirrato; 4C in metano; 6C in CO2; il bilancio dell’ATP è di 16 moli (3,2 per mole di esoso fermentato).
Una particolare fermentazione è quella che si sviluppa per opera dei batteri lattici. Poichè la loro attività si sviluppa in maniera tanto maggiore quan to minore è il pH e poiché il loro prodotto finale (acido lattico) è un acido organico molto più forte degli AGV (pKa 3,86), se le razioni sono ricche di zuccheri solubili, l’azione dei lattici è esaltata, il pH è influenzato dalla loro presenza e se non si
rimuove la causa (uso dei tamponi, riduzione dell’apporto di zuccheri semplici), l’acido lattico si accumula nel rumine causando la riduzione dell’attività degli altri batteri fino alla loro scomparsa. Questa descritta è l’eziologia della acidosi lattica, una forma di disturbo digestivo fastidiosa per i ruminanti.
Le fermentazioni sui substrati azotati
Le proteine ed i composti azotati non proteici (NPN) contenuti nella razione vengono modificati dalla micropopolazione ruminale. Questa degrada più facilmente le proteine solubili nel liquido ruminale e l’NPN rispetto alle proteine insolubili. Le proteine degradabili (solubili ed insolubili) e l’ NPN danno origine ad un pool di azoto fermentescibile che, attraverso prodotti intermedi (ammoniaca, peptidi e aminoacidi), è utilizzato dai microorganismi per la sintesi delle proteine cellulari (proteina grezza microbica). L’azoto ammoniacale, in presenza di ATP derivante dalla fermentazione dei carboidrati, è organicato dai batteri (prevalentemente dai cellulosolitici) e trasformato in proteine: la possibilità di
organicazione dell’azoto, consente ai ruminanti di fornirci alimenti di alto valore biologico (latte e carne) a partire da composti azotati di basso valore biologico (sostanze azotate vegetali) o addirittura a partire dall’azoto inorganico.
Se l’NH 4 fosse in eccesso rispetto alla disponibilità di ATP (oppure l’ATP fosse in difetto rispetto alla disponibilità azotata), la quota ammoniacale in ecc esso filtra la parete del rumine, arriva al fegato in cui è trasformata in urea (l’ammoniaca è tossica) che viene eliminata per via renale oppure riciclata per via salivare; il riciclo dell’urea per via salivare consente un risparmio di azoto importante per animali che si alimentano con foraggi molto poveri in sostanza azotate (es.paglie).
Le proteine alimentari non degradate nel rumine (escape ruminale proteico)+ le proteine dei batteri che colonizzano le digesta o il liquido ruminale che oltrepassano l’ostio reticolo-omasale, rappresentano l’entità di proteine realmente a disposizione dell’ animale per la digestione successiva.
2.4 L’INGESTIONE ALIMENTARE
L’ingestione alimentare è la quantità (espressa in s. s.) di un determinato alimento (o razione) che un determinato animale è in grado di assumere ad libitum nell’arco di 24 ore. Se lo esprimiamo in termini assoluti (kg di s.s.) e si denomina ingestione, invece in termini relativi (in % del peso corporeo dell’animale), prende il nome di livello di ingestione, o in termini relativi ad un determinato alimento di riferimento (normalmente l’erba) e allora la chiamiamo capacità di ingestione.
In tutti gli animali (ruminanti e non) l’ingestione alimentare dipende:
- dall’animale (specie, mole, stato d’ingrassamento , gestazione);
- dall’alimento (composizione chimica, aspetto fisico, appetibilità);
- dalle modalità di distribuzione e presentazione ( management);
- dal clima;
- dalla conduzione dell’allevamento (presenza d’alt ri animali, sanità, ecc.).
L’ingestione è regolata dai centri della sazietà e da quelli della fame, collocati nell’ipotalamo. Questi centri agiscono come un interruttore: quando il centro è orientato verso la fame il comportamento dell’animale è orientato verso la ricerca e l’ingestione dell’alimento e, viceversa. Il centro è influenzato da recettori meccanici che si trovano nelle pareti del rumine che, percependo lo stato di replezione dell’organo, intervengono con l’interruzione o l’in izio dell’alimentazione, e da recettori metabolici, collocati nel fegato o nell’intestino, che monitorano l’entità di metaboliti in circolazione. L’ampiez za dei segnali di fame/sazietà (Is) è condizionato dal grado di distensione del rumine (S), dalla concentrazione ematica dei metaboliti (C), dalla numerosità dei recettori fisici (Ns) e chimici (Nc) secondo la relazione studiata da Conrad e altri:
Is = (S x Ns) ± (C x Nc).
La digeribilità degli alimenti influenza la loro in gestione in quanto tanto più rapidamente un alimento è degradato nel rumine, tanto più rapidamente scompare per lasciare il posto ad un nuovo alimento. Con alimenti a bassa digeribilità prevalgono gli stimol i legati al grado di replezione ruminale, con il crescere della digeribilità, entrano in gioco i vin coli all’ingestione di tipo metabolica. La digeribilità delle razioni è legata al contenuto in pareti cellulari ed in proteina, ed esiste una relazione inversa fra ingestione e contenuto in CS della razione ed un rapporto diretto fra ingestione e concentrazione proteica. Nel primo caso l’aumento del contenuto in fibra riduce la digeribilità e aumenta i MRT; nel secondo caso, l’aumento di disponibilità azotata co nsente ai micoorganismi di riprodursi con maggiore efficienza e quindi di attaccare la fibra con superiore efficacia. In entrambi i casi la velocità di
svuotamento del rumine aumenta con conseguente più frequente rilascio dei recettori dello stato di distensione delle pareti e quindi più frequente segnale di fame all’ipotalamo.
2.5 IL COMPORTAMENTO ALIMENTARE DEI POLIGASTRICI
La più importante differenza fra le diverse specie ruminanti deriva dal rapporto allometrico fra apparato digerente e massa corporea e fabbisogni energetici e massa corporea: il primo mostra un esponente per il peso corporeo di 0,94, o di 1,0, mentre per il secondo tale esponente è 0,75.
Ciò in pratica significa che:
- i piccoli ruminanti (PR) hanno una massa corporea 10-12 volte inferiore a quella dei grandi ruminanti (GR).
- ambedue presentano un volume totale dell’apparato digerente che varia in maniera proporzionale al peso corporeo e che mediamente rappresenta il 13-18% di questo.
- i fabbisogni nutritivi di mantenimento ( metabolismo basale + attività spontanee ) sono meno che proporzionali al peso corporeo essendo, infatti, legati al cosiddetto peso metabolico (PM = peso corporeo0,75), per cui i grandi ruminanti tendono ad avere un maggiore volume di apparato digerente per unità di fabbisogni energetici di mantenimento che non i piccoli ruminanti. Per compensare la bassa capacità digestiva i piccoli ruminanti aumentano la velocità di transito degli alimenti nel rumine ed hanno livelli di ingestione più elevati dei grandi ruminanti; ciò comporta una riduzione dei tempi di
ritenzione ruminale ed una più bassa digeribilità d elle frazioni a lenta fermentazione (soprattutto fibra), compensata da maggiori ingestioni giornaliere di sostanze digeribili.
Altre differenze anatomiche riguardano:
- il rapporto fra volume ruminale e volume dell’apparato digerente nel suo complesso. I GR hanno un intestino tenue relativamente più piccolo dei PR e digeriscono con minore efficienza gli amidi che possono uscire dal rumine non fermentati. Gli ovini e caprini hanno una efficienza nella digestione degli amidi superiore a quella dei bovini per una maggiore quota di intestino in cui può avere sede la digestione enzimatica;
- la dimensione delle ghiandole parotidi è molto maggiore nei PR rispetto ai grandi in quanto nei primi hanno anche la funzione di produrre sostanza in grado di tamponare gli alcaloidi presenti nei foraggi di cui si alimentano (fronde di alberi e di cespugli);
- la mobilità del labbro superiore di PR rispetto ai GR e la possibilità dei primi di esplorare con più efficacia lo spazio tridimensionale.
Le conseguenze di queste differenza antomo-fisiologica fra PR e GR sono che i primi rispetto ai secondi:
a) devono avere livelli di ingestione più alti per soddisfare i loro fabbisogni di mantenimento cui consegue una maggiore velocità di transito ed una m inore digeribilità della fibra;
b) tendono ad essere più selettivi;
c) sono più influenzati nell’ingestione alimentare dalla quantità e dalla dimensione della fibra degli alimenti;
d) devono spendere più tempo per ingerire e ruminare ciascun kg di alimento;
e) tendono ad avere una maggiore digeribilità delle granelle e delle razioni ricche d’energia.
Sulla base di queste differenze anatomo-fisiologiche, i ruminanti sono stati classificati in 3 categorie:
- i concentrate selectors, a cui appartengono le specie che vivono nei boschi (cervi, daini, ecc..) e che selezionano diete molto ricche in principi nutritivi e povere in fibra, ma con elevate concentrazioni di alcaloidi;
- i grazers, a cui appartengono i bovini e i bufalini che pascolano esclusivamente (o quasi) le erbe
e che mal si adattano ad utilizzare le fronde degli alberi e dei cespugli;
3) gli intermediate feeders, a cui appartengono i caprini e gli ovini che hanno un comportamento intermedio, ma più simile ai selectors i caprini e ai grazers gli ovini.
2.6 CENNI DI METABOLISMO (con particolare riguardo ai poligastrici)
Il metabolismo energetico degli animali erbivori differisce da quello degli onnivori (e dei granivori quali gli avicoli) in quanto i nutritivi che derivano dalla digestione sono costituiti nel primo caso prevalentemente da acidi grassi volatili e nel secondo da glucosio (e lipidi).
La differenza fondamentale fra poligastrici (e parzialmente anche erbivori monogastrici e ciecotrofi) e monogastrici (onnivori e granivori) sta nel fatto che nei primi i polisaccaridi strutturali ( CS) e quelli di riserva (CNS) sono degradati prevalentemente nel rumine con la produzione dei 3 AGV, mentre nei secondi il glucosio deriva dalla digestione intestinale degli amidi. Nei ruminanti il metabolismo energetico finale è sostenuto direttamente dall’acetato e dal betaidrossi-butirrato d’origine ruminale, che entra nel ciclo Krebs direttamente, mentre nei monogastrici tale funzione è assolta dal glucosio che produce acetato (in realtà acetil_COA) attraverso l a glicolisi.
I poligastrici, per i quali soltanto una quota limitata di glucosio proviene dalla digestione degli amidi, sintetizzano glucosio nel fegato a partire principalmente da propionato, ma anche dagli aminoacidi neoglucogenetici e dal glicerolo che deriva dall’idrolisi dei lipidi.
Possiamo distinguere il metabolismo in assorbitivo e in postassorbitivo: nel primo, successivamente ai pasti, prevalgono i processi anabolici in quanto i nutritivi assorbiti a livello ruminale e intestinale sono in elevata concentrazione; nel secondo, lontano dai pasti, prevalgono processi catabolici.
Nei ruminanti è difficile fare questa distinzione in quanto la continua messa d’alimenti in fermentazione nel rumine assicura un continuo rifornimento di metaboliti; tuttavia, in alcune circostanze la domanda metabolica dovuta alla produzione (ad es. animali con elevata produzione di latte; fase finale della gestazione), può essere ec cedente rispetto all’apporto di nutritivi con la razione alimentare assunta dagli animali. Parleremo pertanto di bilancio metabolico positivo (in cui gli apporti dal digerente superano la domanda metabolica per il mantenimento e la produzione) e di bilancio metabolico negativo (in caso contrario).
2.6.1 Bilancio metabolico positivo
In questo stato i flussi sono orientati dall’apparato digerente al fegato e dal fegato ai tessuti periferici. I principali metaboliti derivanti dalla digestione (aminoacidi, acetato-butirrato, propionato) sono trasferiti dalla vena porta al fegato.
L’acetato non è trattenuto dal fegato e passa nella circolazione generale. Viene captato dai tessuti periferici e inserito direttamente nel ciclo di Krebs per la produzione di energia. Può anche essere utilizzato dal tessuto adiposo per la produzione di acidi grassi attraverso la beta-condensazione o il
ciclo del malonato. Gli acidi grassi sono poi esterificati con glicerina e depositati sotto forma di lipidi nel tessuto stesso.
Il butirrato (in realtà l’ac. butirrico filtra la p arete del rumine ed è trasformato in beta-idrossi butirrato) segue lo stesso destino in quanto nei tessuti è idrolizzato ad acetato.
Il propionato è trattenuto dal fegato che, attraverso la gluconeogenesi, lo converte in glucosio. Questo metabolita è rilasciato dal fegato ed è impiegato per la nutrizione del tessuto nervoso, per la produzione del latte e per la nutrizione del feto. E’ indispensabile per la sintesi dei grassi nel tessuto adiposo in quanto fornisce la glicerina necessaria per l’esterificazione degli acidi grassi. Il glucosio è usato per fornire il substrato energetico per la contrazione muscolare anaerobia. Può essere, in piccola parte, accumulato nel fegato e nei muscoli sotto forma di glicogeno; l’eventuale quota eccedente è convertita in grasso nel tessuto adiposo o negli adipociti degli altri tessuti.
Gli aminoacidi subiscono nel fegato la transaminazione per la sintesi degli AA non essenziali (quelli essenziali sono apportati dalla razione); nel caso dei ruminanti il concetto di AA essenziale è vago, perchè il valore biologico delle proteine di cui ècomposta la micropopolazione ruminale è elevato, per cui questi animali non hanno vere esigenze in AA. Gli AA rilasciati dal fegato sono captati dai tessuti per il turnover proteico, per la deposizione delle proteine (durante l’accrescimento) e per la produzione (ad es. proteine del latte). Una quota di AA può es sere impiegata nel fegato per la sintesi di glucosio (AA glucogenetici).
2.6.2 Bilancio metabolico negativo
In questo stato i flussi sono orientati dai tessuti (eccetto il nervoso) al fegato.Si richiede la sintesi di glucosio: uno sbilanciamento energetico significa, pertanto, una carenza metabolica di glucosio e l’intero organismo è impegnato a tamponare questa domanda.
La domanda iniziale di glucosio può essere soddisfa tta con la riconversione del glicogeno, che era stato depositato nel fegato e nei muscoli. L’entità di gl icogeno così disponibile è però molto bassa ( tampo na i casi di grave emergenza); per la domanda energetica l’ organismo si rivolge al tessuto adiposo.
L’acetato e gli acidi grassi a più lunga catena non possono essere utilizzati per produrre glucosio. Tutti i composti C3 (acido propionico, glicerolo, ecc.) possono essere utilizzati per la gluconeogenesi; invece i composti C2 o i loro polimeri no. Il risultato è che dall’idrolilsi dei trigliceridi del tessuto adiposo può essere utilizzata per la produzione di glucosio soltanto la glicerina. La forte richiesta di energia (glucosio) porta però alla mobilizzazione degli AG dal tessuto adiposo: sono captati dai mitocondri per la produzione di ATP e, la quota in eccesso, è eliminata dal fegato sotto forma di corpi chetonici.
Se la richiesta di glucosio è imponente e il rifornimento alimentare di propionato insufficiente (razioni povere in amidi; livello di ingestione basso), il fegato inizia a produrre una quantità notevole di co rpi chetonici che sono utilizzati dai tessuti per fini energetici, ma che sono anche eliminati dall’animale con l’expirium e con le urine; questa perdite netta di energia aumenta lo sbilanciamento energetico e, se somministriamo razioni più adeguate, possiamo andare in contro a CHETOSI; che è più frequente:
a) nella vacca da latte nelle prime settimane di lattazione, per la nota disincronia fra il massimo livello produttivo (che si verifica intorno alle 6 settimane), ed il massimo livello di ingestione (che si verifica intorno ai 3 mesi);
b) nelle pecore, nelle ultime settimane di gestazione soprattutto con gravidanze plurime, quando la domanda di glucosio del feto è alta e l’ingestione alimentare è limitata proprio dalla presenza del feto.
In sintesi: un bilancio nutritivo positivo da la garanzia che l’energia in eccesso sia conservata sotto forma di lipidi; uno negativo deve comunque assicurare una produzione di glucosio tale da far fronte alla domanda delle funzioni vitali e produttive degli animali ed evitare un aumento dei corpi chetonici circolanti.
2.6.3 Il ciclo dell’urea
L’ammoniaca prodotta in eccesso nel rumine nel corso delle fermentazioni (e lo ione ammonio derivante dalla deaminazione degli AA) è rimossa dal sangue dal fegato e trasformata in urea. Tale funzione epatica è indispensabile in quanto l’NH3 è tossica, soprattutto per il tessuto nervoso in quanto blocca l’ossalacetato che è indispensabile per la respirazione.
Il ciclo dell’urea si svolge in parte entro il mitocondrio dell’epatocita, in parte nel citoplasma; Dei 2 gruppi NH3 dell’urea, uno deriva dal glutammato ed uno dall’azoto ammoniacale, che deve essere smaltito. Quindi per ogni mole di ammoniaca da smaltire una mole di gruppi amminici è di origine endogena. Tutta la reazione è fortemente endoergonica: si consumano 4 moli di legami P per mole di urea sintetizzata.
In sintesi, nei casi in cui si abbia uno sbilanciamento fra apporti energetici e apporti proteici, o nel caso di forte domanda di glucosio che comporta una elevata attività di deamminazione a carico degli AA neoglucogenetici, l’ammoniaca prodotta nel rumine è convertita in urea dal fegato con un aggravio energetico notevole. S pensi che 100 g di proteina grezza inutilizzata nel rumine sono equivalenti per una pecora alla spesa energetica necessaria alla produzione di 200 g di latte.
Fonte: http://agrariafree.altervista.org/appunti%20agraria/zootecnica%20generale/APPUNTIzootecnica%20generale_.pdf
Sito web da visitare: http://agrariafree.altervista.org
Autore del testo: non indicato nel documento di origine
Parola chiave google : Zootecnica generale appunti tipo file : doc
Zootecnica generale appunti
Visita la nostra pagina principale
Zootecnica generale appunti
Termini d' uso e privacy