Comportamenti fisico meccanici del legno
Comportamenti fisico meccanici del legno
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Il comportamento fisico-meccanico del legno
Dispensa integrativa ad uso degli studenti del Corso di Laurea in Disegno Industriale
Il legno, materiale naturale di origine biologica
In questa sede non è ovviamente possibile sviluppare una trattazione completa del “materiale legno”, nei suoi molteplici ed affascinanti aspetti, per i quali si rinvia ai testi fondamentali citati in bibliografia.
D’altra parte riteniamo opportuno richiamarne, sia pure soltanto attraverso una sintetica carrellata terminologica, alcuni aspetti fondamentali che comunque non possono essere ignorati da chi voglia occuparsi non superficialmente della conservazione e del restauro dei supporti lignei.
Variabilità
Anzitutto non si deve mai trascurare che il legno è un materiale naturale, di origine biologica, formatosi nell’ambito di organismi viventi; le sue caratteristiche pertanto non costanti né controllabili in sede di “produzione”, se non attraverso una accurata selezione dei singoli pezzi, e la sua variabilità è notevolissima addirittura all’interno di uno stesso pezzo.
Specie legnose
Il legno viene prodotto da alberi che appartengono a numerose specie (meglio evitare il francesismo “essenza”, che in italiano ha significati differenti). Ogni specie è caratterizzata da un nome botanico (che va scritto in carattere corsivo), formato da due termini latini (indicanti il genere e la specie) e dal nome abbreviato dello studioso che per primo l’ha identificata come entità botanica distinta: p.es. Abies alba Mill. = Abete bianco, Populus alba L. = Gattice (Pioppo bianco), Quercus petraea Liebl. = Rovere, etc. (v. la Tab. 1). A tutt’oggi sono state descritte migliaia di specie legnose, delle quali soltanto qualche centinaio fornisce legname da lavoro che abbia avuto od abbia interesse in Italia, e soltanto qualche decina è stata usata non occasionalmente per la costruzione di supporti lignei di dipinti su tavola. Le norme UNI 2853 - 2854 - 3917 indicano i nomi normalizzati delle specie (o dei gruppi commerciali) che rispettivamente vegetano spontanee in Italia, vi sono state introdotte e coltivate, oppure che non vegetano in Italia ma il cui legname vi viene importato. Raramente il nome unificato fa riferimento ad una entità geografica (p.es. “Ontano napoletano”, “Pioppo canadese”, “Tuia del Pacifico”); in tal caso esso può indicare in qualche modo la zona di origine o di iniziale diffusione della specie, ma non la provenienza effettiva di un determinato segato, tronco o lotto di legname.
Oltre a quelli unificati, sono talora in uso in certe zone od in certi ambienti professionali altri nomi (p.es. “Pino cembro” invece di Cirmolo, “Acacia” invece di Robinia, “Pino di Svezia” invece di Pino silvestre), che ove possibile sarebbe opportuno evitare.
Esistono infine denominazioni più o meno di fantasia, che purtroppo vengono usate spesso nel commercio e che sembrano indicare o garantire (anche se per lo più non è vero) la provenienza del legname . Per quanto l’uso dei nomi normalizzati non sia obbligatorio, esso è certamente raccomandabile, allo scopo di evitare errori e malintesi che sovente possono costituire vere e proprie scorrettezze; al di là degli aspetti più strettamente culturali o commerciali, resta poi il fatto che i legnami ricavati da specie legnose diverse presentano differenze che non si limitano ai caratteri puramente botanici (come ad esempio la morfologia di foglie e fiori) delle specie da cui provengono, ma riguardano anche numerosi ed importanti aspetti tecnici e tecnologici, fra cui ad esempio il colore, l’aspetto, la densità, la stabilità dimensionale, la durabilità naturale, l’omogeneità della struttura, la lavorabilità, l’attitudine alla finitura, le caratteristiche meccaniche, etc., e che
Tab. 1 - Nomi italiani, nomi botanici, valori tipici (in neretto) ed intervallo di variazione della densità, per alcuni legni comunemente impiegati in Italia (da: Giordano, 1988)
NOME ITALIANO |
NOME BOTANICO |
Densità a umidità normale (U=12%) |
Latifoglie |
|
|
Acero, Acero montano |
Acer pseudoplatanus L., Acer sp.p. |
0,58 0,66 0,74 |
Bosso |
Buxus sempervirens L. |
0,94 1,00 1,03 |
Castagno |
Castanea sativa Mill. |
0,37 0,58 0,70 |
Ciliegio |
Prunus avium L. |
0,55 0,62 0,80 |
Faggio |
Fagus sylvatica L. |
0,52 0,73 0,93 |
Farnia |
Quercus robur L. |
0,50 0,82 0,97 |
Frassino |
Fraxinus excelsior L. |
0,52 0,72 0,87 |
Gattice (= Pioppo bianco) |
Populus alba L. |
0,43 0,48 0,53 |
Noce |
Juglans regia L. |
0,63 0,72 0,75 |
Olmo |
Ulmus montana Stokes |
0,46 0,62 0,70 |
Pero |
Pyrus communis L. |
0,50 0,68 0,80 |
Pioppi euramericani |
Populus euramericana (Dode) Guinier |
0,26 0,34 0,52 |
Salice |
Salix alba L., Salix sp.p. |
0,32 0,45 0,55 |
Tiglio |
Tilia cordata Mill., Tilia sp.p. |
0,58 0,65 0,70 |
Conifere |
|
|
Abete bianco |
Abies alba Mill. |
0,31 0,44 0,61 |
Abete rosso |
Picea abies Karst. |
0,30 0,45 0,62 |
Cipresso |
Cupressus sempervirens L. |
0,42 0,61 0,84 |
Larice |
Larix decidua Mill. |
0,38 0,65 0,93 |
Cirmolo (= Pino cembro) |
Pinus cembra L. |
0,32 0,45 0,65 |
Pino silvestre |
Pinus sylvestris L. |
0,38 0,55 0,66 |
quindi occorre accertarsi non soltanto che un nome sia scelto e scritto correttamente, ma che il legno di cui si tratta appartenga realmente alla specie dichiarata .
Durezza, durame, durabilità
Le specie legnose possono essere suddivise in Conifere (dette anche - meno prpriamente - resinose od aghifoglie: p.es. Abete bianco, Abete rosso, Cipresso, Cedro) ed in Latifoglie (p.es. Rovere, Pioppo, Castagno, Noce). A questo riguardo una certa confusione può essere causata dai corrispondenti termini in lingua inglese, dove softwood (letteralmente "legno tenero") significa "Conifera", mentre hardwood (letteralmente "legno duro") significa "Latifoglia"; si tratta quindi di una suddivisione puramente botanica, che nulla ha a che fare con la durezza del legno, che invece ne è una proprietà meccanica, misurabile con apposite prove di laboratorio (metodi Janka, Chalais-Meudon, Brinnell, etc.).
Un'altra importante proprietà del legno è la sua durabilità (naturale, oppure conferita mediante idonei trattamenti preventivi), che esprime la capacità potenziale di un legname a resistere agli organismi xilofagi quali i funghi (che possono provocare nel legno alterazioni cromatiche ed anche marciume, sinonimo di carie), o gli insetti (che possono scavare nel legno gallerie, fino a ridurlo in polvere, o meglio in rosume); la effettiva durata in opera di un manufatto ligneo dipende non soltanto dalla sua durabilità, ma anche dalle condizioni (soprattutto ma non esclusivamente quelle termoigrometriche) dell'ambiente in cui si trova. Nonostante l'assonanza dei termini, la durabilità del legno nulla ha a che fare con la sua durezza (p.es. il Faggio è piuttosto duro, ma non è affatto durabile, e viene facilmente attaccato sia dai funghi che da insetti).
La parte più durabile di un tronco è generalmente il durame (talvolta detto anche legno perfetto oppure cuore), che ne costituisce la parte interna dove le cellule costituenti i tessuti legnosi non partecipano più ai processi vitali dell'albero, e svolgono essenzialmente funzioni di sostegno. In alcune specie le pareti cellulari delle cellule costituenti il durame sono coperte od impregnate di estrattivi (tannini, fenoli, etc.), che conferisco al legno una durabilità naturale più o meno elevata, a seconda della loro natura e quantità; se gli estrattivi conferiscono al durame una colorazione (più o meno intensa), esso risulta differenziato rispetto all'alburno, zona anulare più chiara (albus = bianco) nella quale le cellule parenchimatiche sono ancora vive finché l'albero è vivo, e contengono quindi sostanze di riserva (zuccheri, amidi, etc.); di conseguenza l'alburno deve sempre essere considerato non durabile, anche se appartiene a specie con durame durabile (p.es. Rovere, Castagno, Cipresso, Cedro, Larice).
Conoscere la struttura del legno per comprenderne il comportamento
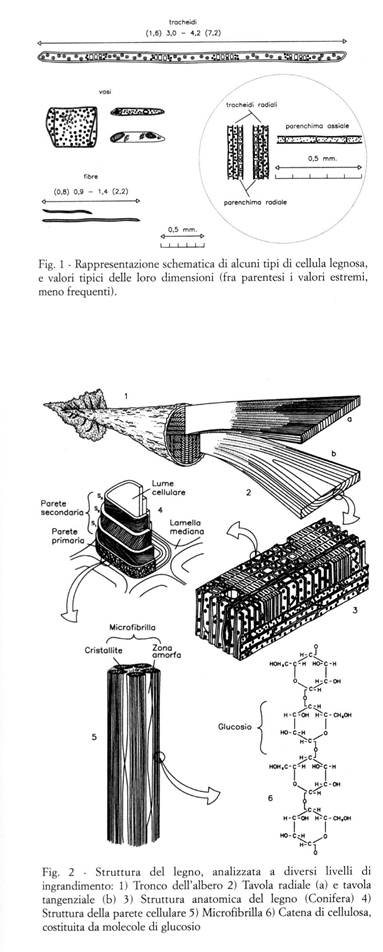
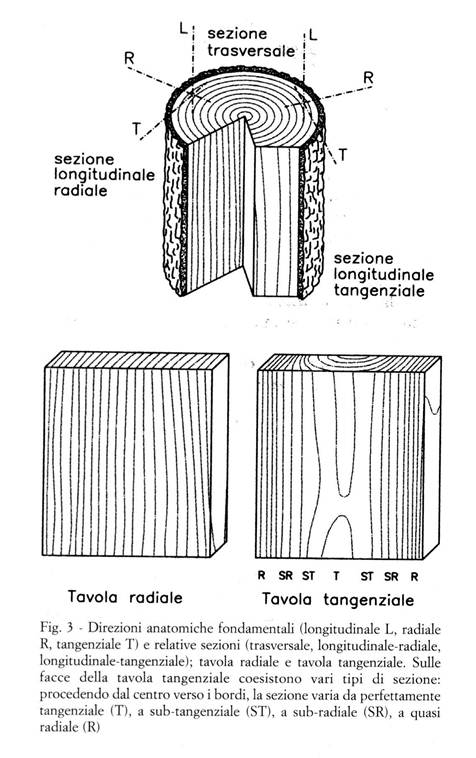
Il legno, o xilema, è prodotto dagli alberi, che sono organismi viventi, ed è costituito da cellule, che già nell'albero in piedi sono per la maggior parte morte e lignificate; esso adempie principalmente alle funzioni di sostegno meccanico, di conduzione idrica, di accumulo di sostanze di riserva, di secrezione di sostanze utili alla vita dell'albero. Le cellule legnose hanno forma allungata, con direzione longitudinale circa parallela all'asse del tronco, e sono costituite da una parete cellulare, che circonda una cavità detta lume cellulare (Fig. 1). La maggior parte delle proprietà fisiche e meccaniche del legno è determinata dalla sua struttura, che può essere analizzata a diversi livelli di ingrandimento (Fig. 2):
- la struttura chimica (elementare e complessa)
- la ultrastruttura della parete cellulare, che può essere osservata con microscopi molto potenti, come ad esempio il TEM (microscopio elettronico a trasmissione)
- la microstruttura dei tessuti legnosi, che può essere osservata con microscopi ottici, o talora con semplici lenti
- la macrostruttura, che può essere osservata ad occhio nudo, o con lenti a modesto ingrandimento.
Struttura del tronco e del legno (macrostruttura), anisotropia
Come è ben noto, l'accrescimento degli alberi avviene mediante la formazione di strati di accrescimento, che nelle regioni a clima temperato ( ) avviene con ciclicità annuale. Tali strati annuali vengono generati dal cambio, sottile strato di cellule situato fra legno e corteccia; durante la stagione vegetativa le cellule del cambio si suddividono e si moltiplicano tangenzialmente e radialmente, generando verso l'interno il legno vero e proprio (xilema, costituito da cellule che muoiono e lignificano) e verso l'esterno altri tessuti essenziali per la vita e la protezione dell'albero (floema, corteccia).
Sulle testate dei tronchi tali strati danno luogo ai ben noti anelli (annuali) di accrescimento, più o meno visibili a seconda della specie legnosa e dello stato della superficie. In genere gli anelli (e più generalmente la venatura del legno, determinata dall'intersezione degli strati di accrescimento con una qualsiasi superficie del legno) sono visibili grazie all'alternanza fra il legno primaverile (meno denso e più chiaro) ed il legno tardivo (più denso e più scuro); questa non è però una regola assoluta, dato che in qualche caso risulta difficile se non impossibile distinguere ad occhio nudo il legno primaverile dal legno tardivo.
Un'altra importante caratteristica del legno osservabile a occhio nudo è il colore, che principalmente dipende dalla eventuale presenza di estrattivi. Oltre che sul colore, gli estrattivi possono influire notevolmente anche su altre proprietà del legno, fra cui la durabilità naturale, la stabilità dimensionale, la permeabilità, la facilità all'essiccazione, l'acidità, la facilità ad essere verniciato od incollato, la tendenza a corrodere le ferramenta e gli utensili in condizioni di elevata umidità, etc.
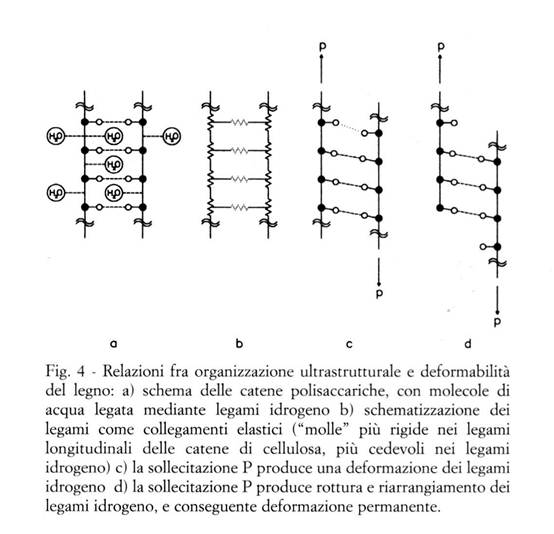
La struttura anatomica del legno ne definisce le tre direzioni anatomiche fondamentali (Fig. 3):
- la direzione longitudinale (direzione della fibratura), definita dalla direzione complessiva degli assi delle cellule legnose
- la direzione radiale, corrispondente a quella dei raggi parenchimatici
- la direzione tangenziale, tangente agli strati di accrescimento.
Ammettendo che il tronco degli alberi sia cilindrico (il che è solo in parte vero, dato che i tronchi possono essere più o meno rastremati, curvi, a sezione non circolare, etc.), le tre direzioni anatomiche fondamentali sono in ogni punto mutuamente perpendicolari; tipicamente la direzione longitudinale (o assiale) L corrisponde a quella dell'asse e delle generatrici del tronco, mentre la radiale R e la tangenziale T corrispondono rispettivamente ai raggi ed alle tangenti delle sezioni circolari rette.
In ogni punto, le tre direzioni anatomiche definiscono poi tre sezioni fondamentali: trasversale TR, longitudinale-radiale (o semplicemente: radiale) LR, e longitudinale-tangenziale (o semplicemente: tangenziale) LT.
Dato che per punti diversi di un stesso pezzo di legno le direzioni anatomiche fondamentali R e T possono essere diverse tra loro (la direzione L può esserlo se il pezzo presenta deviazioni della fibratura), su una stessa superficie di un pezzo di legno possono coesistere sezioni diverse.
Queste considerazioni sono essenziali per comprendere il comportamento anisotropo del legno. Tale comportamento si manifesta per molte proprietà, ed in particolare per ritiri e rigonfiamenti, caratteristiche meccaniche, permeabilità ai liquidi ed ai gas.
Microstruttura
La maggior parte delle cellule legnose ha forma allungata ed è disposta circa parallelamente all'asse del fusto. Nelle Conifere tali cellule sono chiamate tracheidi, e svolgono funzioni sia di sostegno meccanico che di conduzione della linfa; nelle Latifoglie le due funzioni vengono assolte da cellule specializzate, le fibre (che svolgono funzioni di sostegno) ed i vasi (che svolgono funzioni di conduzione).
In tutte le Conifere e Latifoglie esistono poi cellule parenchimatiche (dove si accumulano periodicamente le sostanze di riserva necessarie all'accrescimento dell'albero: amidi, zuccheri, etc.) organizzate in raggi parenchimatici: strutture simili a nastri, che decorrono in direzione radiale dal midollo fino al cambio, e la cui altezza è orientata secondo la direzione longitudinale delle cellule di sostegno. Quando sono costituiti da un solo strato di cellule (raggi monoseriati) oppure sono pluriseriati ma costituiti da pochi strati, i raggi parenchimatici non risultano visibili ad occhio nudo; quando il loro spessore è costituito da molti strati di cellule, risultano più o meno facilmente visibili ad occhio nudo.
In qualche specie sono presenti, con maggiore o minore abbondanza, anche cellule parenchimatiche longitudinali, ed altri tipi di cellule (come ad esempio quelle formanti i canali resiniferi), atte a secernere resine, gomme o lattici.
La presenza di cellule di vario tipo e la loro disposizione, congiuntamente alla presenza di altri caratteri anatomici, possono essere utilizzate per la identificazione (macroscopica o microscopica) della specie legnosa, che peraltro richiede in generale conoscenze specialistiche e notevole esperienza.
Ultrastruttura della parete, e sua influenza sulle caratteristiche del legno
La conoscenza almeno sommaria della struttura e della ultrastruttura del legno costituisce uno strumento indispensabile per comprenderne il comportamento fisico e meccanico, soprattutto per quanto riguarda l'igroscopicità e l'anisotropia (comportamento diverso nelle diverse direzioni anatomiche L,R,T). L’anisotropia in particolare si manifesta a diversi livelli di organizzazione strutturale del legno, dalle modalità aggregazione dei singoli costituenti chimici fino alla struttura dei tessuti.
I costituenti del legno
I costituenti chimici del legno sono rappresentati dalle tre seguenti principali categorie di composti: le olocellulose (60-70%), le lignine (20-30%) e gli estrattivi (1-15%).
Le olocellulose rivestono un ruolo fondamentale nel determinare il comportamento meccanico e nel regolare gli scambi igroscopici del legno; possono a loro volta essere suddivise in cellulosa, costituita da lunghe catene di glucosio (uno zucchero a 6 atomi di carconio), ed emicellulose composte invece da zuccheri a 5 e 6 atomi di carbonio con un grado di polimerizzazione più ridotto rispetto alla cellulosa.
Le lignine, composti fenolici insolubili, costituiscono una specie di matrice incrostante che contribuisce a rendere più solido il collegamento fra le catene polisaccariche, aumentando la resistenza meccanica del legno e guidandone il comportamento termoplastico.
Gli estrattivi infine comprendono composti di varia natura, dagli amidi fino ai fenoli ed ai derivati dell’isoprene; possono essere estratti dal legno con l’ausilio di solventi polari (acqua, alcool), e nonostante siano presenti in quantità minore rispetto agli altri costituenti, possono conferire al legno importanti proprietà, quali colore, durabilità naturale, stabilità dimensionale, facilità all'incollaggio ed alla finitura, etc.
Organizzazione ultrastrutturale ed anisotropia del comportamento meccanico
A livello di organizzazione ultrastrutturale (Fig. 2) l’unità di riferimento del legno è rappresentata dalle microfibrille, strutture eteropolimeriche costituite nella loro parte centrale (cristallite) dalla aggregazione di singole catene di cellulosa, alla quale si uniscono nella regione periferica (zona amorfa) anche le emicellulose.
Già a questo primo grado di organizzazione del legno, è possibile individuare l’origine di alcuni suoi importanti comportamenti tipici; pertanto, pur senza voler entrare nel dettaglio dei meccanismi che intervengono nella formazione delle microfibrille, appare importante esaminarne più approfonditamente alcuni aspetti.
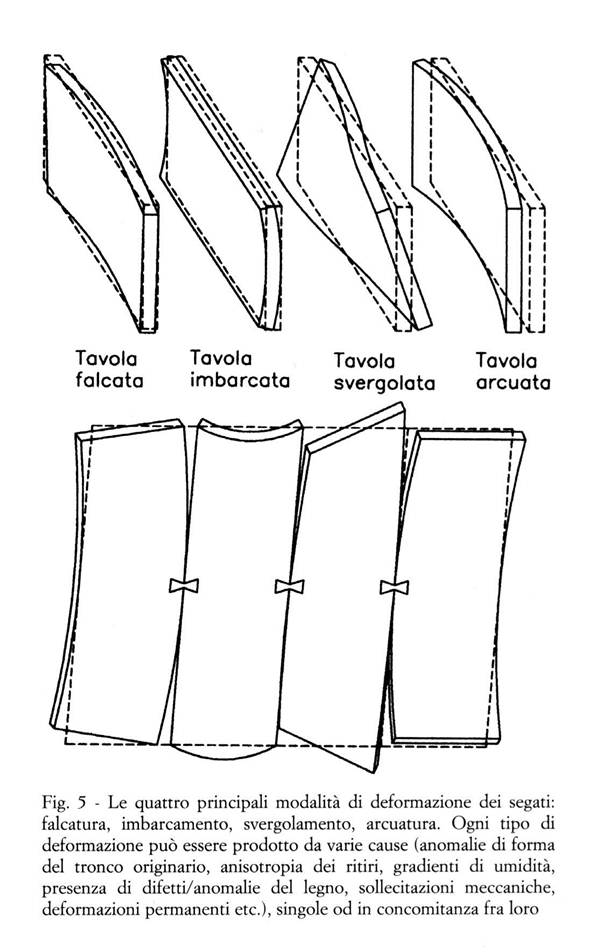
Il legame che permette la congiunzione longitudinale delle singole unità di glucosio è piuttosto stabile (60-80 cal/mole) e non facilmente scindibile; paragonando l’energia di questo legame ad una molla (Fig. 4b), questa può essere considerata piuttosto rigida e capace di conferire alla cellulosa (e quindi alla microfibrilla) una ottima resistenza alle sollecitazioni longitudinali (compressione e trazione). Come si può osservare nella formula di struttura, sulla catena di cellulosa sono presenti numerosi gruppi ossidrilici (R-OH) i quali possono, con altri gruppi simili, formare legami di tipo polare detti legami idrogeno. Rispetto ai precedenti, i legami idrogeno sono caratterizzati da una energia di legame molto più bassa (3-5 cal/mole), tuttavia il grande numero di gruppi ossidrilici presenti sulla molecola di ogni zucchero permette la formazione di un gran numero di legami idrogeno, consentendo alle catene di cellulosa di legarsi fra di loro dando luogo a strutture stabili più complesse quali la microfibrilla. A causa della ridotta energia del legame idrogeno il comportamento delle microfibrille risulta anisotropo, con resistenza piuttosto modesta alle sollecitazioni trasversali (nella Fig. 4b il legame idrogeno è rappresentato da molle più piccole e meno rigide rispetto a quelle usate per il legame longitudinale).
In conclusione, la presenza dei due diversi tipi di legame che collegano longitudinalmente e trasversalmente le catene polisaccariche della microfibrilla, ne spiega il diverso comportamento meccanico nei riguardi di sollecitazioni parallele o trasversali rispetto al suo asse principale.
Organizzazione ultrastrutturale e anisotropia del comportamento igroscopico
Non tutti i gruppi ossidrilici (R-OH) sono però impegnati nella struttura della microfibrilla; nella zona amorfa alcuni di questi rimangono liberi, e fungono da punti di aggancio per altre sostanze in grado di formare legami polari. Fra queste la più importante è sicuramente rappresentata dall’acqua, che grazie alla sua spiccata polarità può formare legami idrogeno con grande facilità. Come esemplificato nella Fig. 4a, l’acqua sfruttando la presenza dei gruppi (-OH) degli zuccheri può legarsi alla struttura della microfibrilla diventando di fatto uno dei costituenti del legno. In realtà la dinamica con cui l’acqua si lega alle microfibrille è più complessa, e non si presta ad essere trattata in questa sede; tuttavia appare opportuno evidenziarne almeno i due seguenti importanti aspetti:
- il numero di gruppi (-OH) che rimangono disponibili all’interno delle microfibrille è molto grande, ma finito, per cui la quantità di acqua che il legno può mantenere direttamente legata alla sua struttura risulta limitata (acqua di saturazione, v. il § 3.2.2);
- la molecola di acqua, se pur molto piccola, possiede una sua dimensione fisica ben definita, per cui a seguito dei fenomeni di equilibramento igroscopico (v. il § 3.2.3), legandosi o liberandosi dalla struttura microfibrillare (adsorbimento o desorbimento) ne provoca un aumento o una riduzione dimensionale (rigonfiamento o ritiro). Inoltre, la disposizione dei gruppi (-OH) lungo le catene (Fig. 2) fa sì che queste variazioni dimensionali interessino piuttosto le direzioni trasversali della microfibrilla che non la direzione longitudinale, determinando in questo modo un'ulteriore anisotropia (quella dei ritiri / rigonfiamenti).
Organizzazione ultrastrutturale e deformabilità del legno
Nella Fig. 4c è schematizzato il comportamento delle catene polisaccariche all’interno della microfibrilla a seguito di una sollecitazione che si mantenga al di sotto del limite di elasticità (v. il § 3.4.1); la conseguente deformazione complessiva della struttura (allungamento od accorciamento, a seconda dei casi) può essere considerata come la risultante del comportamento dei legami longitudinali, più forti (le “molle” più rigide), e dei legami idrogeno, più deboli (le "molle" più cedevoli). L’applicazione della sollecitazione provoca soltanto una deformazione dei legami più forti, mentre nei legami idrogeno può provocare non soltanto deformazioni, ma anche fenomeni di rottura (Fig. 4d); in quest'ultimo caso il gruppo ossidrilico (-OH) che si libera tende a riformare un nuovo legame con un altro ossidrile vicino. L'energia assorbita nel corso di questo processo di rottura e riformazione dei legami compensa il lavoro speso dalla sollecitazione applicata.
Questo fenomeno di scorrimento reciproco delle catene è favorito dalla presenza di molecole di acqua (la resistenza meccanica e la rigidezza del legno diminuiscono con l’aumentare dell’umidità), ed è considerato come una delle spiegazioni più probabili ai fenomeni di scorrimento viscoso manifestati dal legno sotto carico (v. il § 3.4.1).
Organizzazione macrostrutturale
Il comportamento del legno non è sufficientemente spiegato dal suddetto comportamento delle microfibrille; occorre invece esaminare anche il modo in cui queste si organizzano all’interno della parete cellulare e come le singole cellule si dispongono nel tessuto legnoso.
La parete cellulare (Fig. 2) può essere considerata come una struttura poli-lamellare costituita da vari strati, diversi per composizione e struttura. La parete secondaria esercita l’influenza più marcata sul comportamento meccanico complessivo della cellula; a sua volta la parete secondaria è costituita da tre strati, denominati S1, S2, S3 (non sempre quest'ultimo è presente). Ogni strato è costituito dall'aggregazione di fasci di microfibrille, il cui numero può variare in funzione sia dello strato (più numerosi nello S2), sia del periodo e delle condizioni di crescita dell’albero, determinando in tal modo la formazione di cellule con parete di diverso spessore (legno primaverile e tardivo, v il § 2.1). Siccome le microfibrille possiedono un comportamento (meccanico ed igroscopico) spiccatamente anisotropo, il loro arrangiamento all’interno della parete può avere un'influenza determinante sul comportamento della cellula. In condizioni normali l’orientamento delle microfibrille dello strato S2, che quantitativamente è il più importante, risulta circa parallelo all’asse longitudinale della cellula. Tale parallelismo implica di fatto il trasferimento delle proprietà delle microfibrille alla cellula, che infatti risulta spiccatamente anisotropa sia nei riguardi delle sollecitazioni meccaniche, dove manifesta una grandissima resistenza a trazione ed a compressione, sia nei riguardi delle variazioni dimensionali di origine igroscopica, che sono trascurabili longitudinalmente e molto forti trasversalmente (la differenza fra il ritiro radiale e quello tangenziale è dovuta ad altri fattori, fra cui il diverso grado di lignificazione fra pareti radiali e tangenziali, che non è opportuno approfondire in questa sede).
Quanto sopra esposto vale tipicamente per il legno "normale"; nel legno "anomalo" (legno di compressione, o canastro; legno giovanile, v. il § 2.4) le microfibrille tendono a disporsi con un angolo piuttosto inclinato rispetto all'asse longitudinale delle cellule, che presentano allora variazioni dimensionali molto forti anche in direzione longitudinale.
Al successivo livello di organizzazione, cioè a quello del tessuto legnoso costituito dall'insieme delle cellule (Fig. 2), il comportamento del legno rimane coerente con quello delle singole cellule, dato che la maggior parte di esse tende a disporsi con l'asse maggiore parallelo all’asse del tronco, determinando con ciò la direzione della fibratura. Questo spiega l’importanza che deve essere attribuita alle deviazioni della fibratura, che possono determinare nelle tavole comportamenti deformativi anche notevolmente anomali (Giordano, 1981).
Anomalie e difetti
Oltre alla venatura del legno, ad occhio nudo possono essere osservati varie anomalie strutturali (che quando comportano effetti sfavorevoli vengono chiamate difetti), come nodi, deviazioni della fibratura, tasche di resina, cipollature, legno giovanile, legno di reazione, etc. Per motivi di spazio in questa sede non è possibile trattare l'argomento "anomalie e difetti", che pure è molto importante dal punto di vista pratico, perché raramente un pezzo do di legno in dimensione d'impiego potrà essere netto, cioè perfettamente regolare e completamente privo di anomalie.
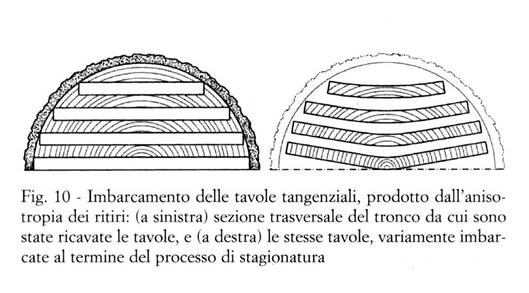
Vale la pena di richiamare che mentre l'imbarcamento del legno è dovuto all'anisotropia dei ritiri trasversali (v. il § 3.3.4), ed è quindi tipicamente presente in tutte le tavole tangenziali, altri tipi di deformazione delle tavole (arcuatura, falcatura, svergolamento; v. la Fig. 5) sono in genere riferibili ai suddetti difetti/anomalie del legno (Giordano, 1981).
Alcune caratteristiche fisico-meccaniche
Essendo il legno un materiale naturale, di origine biologica, con elevata variabilità, non ha senso attribuire valori unici e precisi alle sue caratteristiche quantitative (p.es. attribuire ad una specie legnosa un unico, ben preciso valore di densità o di resistenza meccanica); ed è poco esatto attribuire un valore medio, se non è definito l'insieme di valori da cui la media deriva. Si preferisce quindi far riferimento a valori tipici, e ad intervalli di variazione.
Densità
Dato che le pareti cellulari di qualsiasi specie legnosa sono costituite da cellulosa, emicellulose e lignina, e che tali sostanze hanno densità abbastanza simili ( ), ne consegue che per qualsiasi specie legnosa e per qualsiasi pezzo di legno la densità delle pareti cellulari è circa la stessa, e vale approssimativamente 1,53 g/cm3, o 1530 kg/m3. La diversa densità rilevabile in pezzi appartenenti a specie diverse, oppure in pezzi diversi di una stessa specie, è quindi dovuta alla loro diversa porosità, cioè alla diversa proporzione fra le cavità cellulari e le pareti cellulari esistenti in un dato volume di legno; inoltre per uno stesso pezzo, la densità può cambiare nel tempo al variare dell'umidità del legno, e del suo eventuale degrado (p.es. pirolisi dovuta ad elevate temperature, attacchi di funghi xilofagi, gallerie scavate da insetti); la densità del legno può anche essere alterata dall'eventuale presenza di estrattivi, dal loro dilavamento, oppure dall'impregnazione con sostanze di vario genere (preservanti, consolidanti, adesivi, etc.).
Esistono legni estremamente leggeri (p.es. la Balsa, Ochroma spp., proveniente dall'America centrale, con densità a umidità normale dell'ordine di 50-250 kg/m3 e porosità dell'ordine del 96%) ed estremamente pesanti (p.es. il Guaiaco, Guaiacum officinale L., delle Antille e dell'America centrale, con densità a umidità normale dell'ordine di 1100-1300 kg/m3 e porosità dell'ordine del 12%). Nella Errore. L'origine riferimento non è stata trovata. sono riportati i valori tipici di densità per alcuni legni comunemente impiegati in Italia.
Relazioni legno-acqua
Molte delle peculiarità tecnologiche che distinguono il legno dagli altri materiali derivano dalla sua spiccata affinità per l'acqua. Il legno perfettamente anidro (cioè totalmente privo di umidità) è praticamente inesistente in natura, ed anche in un manufatto ligneo stagionato da lunghissimo tempo è sempre presente una certa quantità di umidità, che dipende principalmente dalle condizioni termoigrometriche dell'ambiente nel quale esso è stato conservato.
L'umidità influisce significativamente su praticamente tutte le caratteristiche tecnologiche del legno (variazioni dimensionali, resistenza meccanica, elasticità, deterioramento, etc.) e condiziona pesantemente la conservazione dei manufatti lignei.
Definizione e determinazione dell'umidità del legno
Si definisce umidità (o contenuto di umidità) U del legno il rapporto (espresso come percentuale) tra la massa dell'acqua contenuta in un qualsiasi pezzo di legno, e la massa anidra di tale pezzo:
(m - mo)
U = -------------- x 100 [%]
mo
essendo:
m la massa del pezzo nelle condizioni di umidità che si intende determinare (stato "tal quale")
mo la massa dello stesso pezzo allo stato anidro (cioè assolutamente privo di umidità), che si può p.es. ottenere mantenendolo in una stufa ventilata a 103 °C fino a peso costante.
L'umidità del legno può anche superare il 100% (ciò p.es. avviene per legno fresco, oppure anche solo parzialmente imbibito) ( ).
Oltre alla diretta applicazione della definizione suddetta (metodo gravimetrico, o per doppia pesata), esistono vari altri metodi per la determinazione dell'umidità del legno ( ). In caso di pezzi di grandi dimensioni, oppure di manufatti che non possono essere introdotti nella stufa ventilata onde evitare di danneggiarli, la determinazione dell'umidità può anche avvenire su frammenti opportunamente prelevati ( ).
Sono in corso di elaborazione norme CEN che verranno recepite in tutti i Paesi dell’Unione Europea, e sostituiranno le norme nazionali.
Fra i più comuni nomi di fantasia, che volendo evitare malintesi non dovrebbero assolutamente essere usati, citiamo qui il “Rovere di Slavonia” (Rovere e Farnia, importate in realtà anche dalla Francia, dalla germania, da vari Paesi dell’Est europeo), ed il “Noce del Tanganica” (Aniegré, che non proviene dall’ex-Tanganika né da zone vicine, ma bensì dall’Africa occidentale, e per di più non appartiene affatto alla famiglia botanica del Noce, le Juglandacee).
In genere, salvo casi particolarissimi, per avere una identificazione realmente affidabile della specie legnosa occorre avvalersi dell’esame microscopico, o dell’esame macroscopico condotto da persona che ne conosca chiaramente metodi e limiti.
E’ bensì vero che bravi falegnami e restauratori conoscono bene i legni dei quali sono esperti; ma è altrettanto vero che i loro settori di competenza riguardano soprattutto proprietà e lavorazioni del legno, cosicché non hanno valido motivo per cercar di differenziare legni apparentemente uguali ed aventi uguali proprietà.
Come regola generale, si raccomanda quindi cautela nell’accettare l’identificazione delle specie legnose richiamate in cataloghi o in letteratura non specialistica, a meno che non vengano esplicitamente citate fonti affidabili, o non venga indicato come si sia pervenuti all’identificazione; fra le varie cause di errata identificazione si citano ad esempio le seguenti:
- l’identificazione della specie legnosa potrebbe essere stata ottenuta soltanto a occhio nudo, da personale non specificamente preparato;
- i “pratici” (falegnami e restauratori) potrebbero non essere stati sufficientemente informati della opportunità che la specie legnosa fosse esattamente identificata; potrebbero quindi avere eseguito erroneamente la loro “identificazione” in base a specie che meglio conoscevano, senza prendere in considerazione che vari legni presentano aspetto e proprietà simili, pur appartenendo a specie diverse (p.es. Pioppo, Tiglio e Salice; Abete bianco e Abete rosso; Pini);
- l’identificazione potrebbe non essere stata eseguita direttamente, ma solo ripresa dalla letteratura, da interviste riportanti opinioni personali, dalla tradizione.
Nelle regioni tropicali la periodicità delle stagioni vegetative può essere diversa da quella annuale.
Si definisce densità (sinonimo di massa volumica) il rapporto fra la massa ed il volume di un determinato corpo (oggetto, provino di laboratorio, etc). Si definisce densità relativa di un materiale il rapporto fra la sua densità e quella di un materiale di riferimento (in genere l'acqua distillata, che alla temperatura di 4 °C ha densità di 1 g/cm3, ossia 1000 kg/m3); oppure (il che è equivalente) il rapporto fra la massa del materiale e la massa di un ugual volume d'acqua. Mentre la densità ha le dimensioni di una massa diviso un volume, la densità relativa è un numero puro, adimensionale; così ad esempio la densità tipica del legno di Pioppo (Populus alba L.) a umidità normale vale 0,43-0,53 g/cm3, ossia 430-530 kg/m3, mentre la sua densità relativa vale 0,43-0,53 senza alcuna unità di misura.
Esempio numerico: un pezzo di legno di Noce appena tagliato pesa 360 grammi, stagionato per alcuni mesi in un ambiente protetto e riscaldato pesa 165 grammi, portato allo stato anidro (mantenendolo per oltre 48 ore in stufa ventilata a 103°C) pesa 150 grammi. Applicando la definizione si avrà:
massa del legno allo stato anidro: mo = 150 grammi
umidità allo stato fresco: Uf = [(360-150)/150]* 100 = 140 %
umidità allo stato stagionato Us = [(165-150)/150]*100 = 10 %
Ad esempio la norma italiana UNI 9091 specifica i seguenti quattro metodi: elettrico (parte 1, del 1987), per pesata (parte 2, del 1987), per distillazione (parte 3, del 1989), igrometrico (parte 4, del 1989). I metodi per pesata, per distillazione ed igrometrico possono essere applicati su provini di legno aventi virtualmente qualsiasi forma e dimensione, compresi schegge e trucioli: qualora i provini siano molto piccoli, ed il loro prelievo non danneggi significativamente il pezzo da cui provengono, tali procedimenti possono considerarsi limitatamente non distruttivi.
In molti casi il prelievo di frammenti può essere eseguito con un utensile carotatore, con un succhiello, o con uno scalpello; in caso di materiale molto frammentato cresce però l'influenza degli scambi di umidità (perdite od acquisti) fra legno ed aria, talora accentuati dalle condizioni atmosferiche o dal riscaldamento prodotto dall'attrito fra legno ed utensile; ciò costituisce una possibile fonte di errore, attribuibile non al metodo ma alla preparazione ed al maneggio dei provini. Occorre poi tenere presente la possibile esistenza di gradienti di umidità, cosicché applicando il metodo gravimetrico sull'intero pezzo ne verrà determinata l'umidità media; applicandolo su frammenti, si determineranno valori di umidità riferibili con precisione soltanto alle zone da cui questi sono stati prelevati.
L'intervallo igroscopico
Nell'albero in piedi l'acqua è presente sia all'interno dei lumi cellulari (acqua libera, o di imbibizione) sia incuneata all'interno delle pareti cellulari (acqua di saturazione, o legata alle pareti); si definisce punto di saturazione delle pareti cellulari (o punto di saturazione, o umidità di saturazione) il valore di umidità del legno in corrispondenza del quale tutta l'acqua di imbibizione è stata eliminata, mentre tutta l'acqua di saturazione è ancora legata alle pareti cellulari.
Il valore del punto di saturazione dipende da vari fattori (specie legnosa, presenza di estrattivi, densità del legno, temperatura, storia pregressa del pezzo, etc.), ed indicativamente può variare tra il 22% ed il 40%, ma per la maggior parte dei fini pratici è sufficiente considerare il punto di saturazione convenzionalmente fissato ad U = 30%.
L'importanza del punto di saturazione consiste nel fatto che esso segna una sorta di soglia al di sotto della quale il legno comincia a presentare sensibili variazioni delle sue proprietà fisico-meccaniche al variare dell'umidità. In altre parole, è soltanto l'acqua di saturazione che influenza significativamente le proprietà del legno: quando la sua umidità scende al di sotto del 30% circa, il legno si riduce di volume, si deforma, può fessurarsi, è più rigido e resistente alle sollecitazioni, mentre non si notano significative differenze (a parte ovviamente il peso) quando lo stesso legno passa dal 200% al 40% di umidità. Per questo motivo è invalso l'uso di chiamare stato fresco qualsiasi valore di umidità maggiore del punto di saturazione.
L'equilibrio igroscopico
Il legno è un materiale igroscopico, cioè la parete cellulare ha la capacità di scambiare la propria acqua di saturazione direttamente con l'atmosfera circostante; quando si trova circondato da aria umida, si ha contemporaneamente una continua evaporazione di molecole d'acqua dal legno nell'aria circostante, ed una continua "cattura" di molecole di vapore da parte della parete cellulare.
Una differenza di velocità tra questi due fenomeni (sempre concomitanti) provoca variazioni dell'umidità del legno (adsorbimento, se questa aumenta; desorbimento se diminuisce), mentre velocità identiche corrispondono ad una situazione di equilibrio igroscopico tra legno ed aria circostante ( ).
Il legno tende sempre ad equilibrarsi con l'umidità relativa UR dell'aria circostante; in prima approssimazione, i valori di equilibrio sono simili per tutte le specie legnose, e dipendono soltanto dalle condizioni termoigrometriche dell'ambiente.
Nella Errore. L'origine riferimento non è stata trovata. vengono riportati alcuni valori di umidità di equilibrio tipiche del legno, in funzione dell'umidità relativa e della temperatura dell'aria.
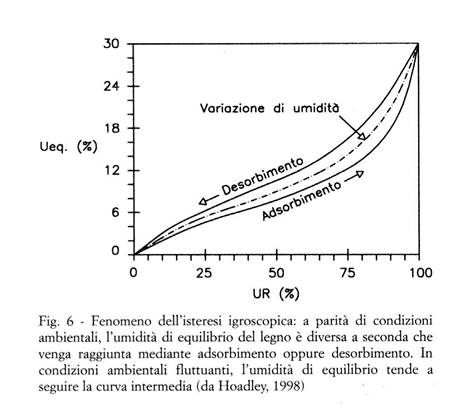
In realtà ciascun pezzo di legno tende ad equilibrarsi a valori che possono essere diversi da quelli tipici, e che dipendono sia da fattori legati al pezzo stesso (specie, caratteristiche proprie del pezzo, estrattivi presenti, etc.) sia del fenomeno dell'isteresi igroscopica, sinteticamente descritto nella Fig. 6.
Qualsiasi pezzo di legno tende a raggiungere o ripristinare l'equilibrio igroscopico con le condizioni termoigrometriche ambientali ogni qual volta queste subiscano una variazione; è pertanto assolutamente errata l'idea, purtroppo ancora diffusa, che "quanto più lunga è la stagionatura, tanto minore sarà l'umidità del legno" (il che implicherebbe che dopo un tempo sufficientemente lungo il legno possa arrivare e mantenersi ad "umidità zero").
L’equilibramento non è un fenomeno istantaneo, ed il tempo necessario al raggiungimento dell'equilibrio igroscopico è relativamente breve soltanto in caso di legno ridotto in sottili trucioli, Tab. 2 - Valori tipici dell'umidità di equilibrio del legno, in funzione dell'umidità relativa e della temperatura dell'aria (da: Giordano, 1981). Con buona approssimazione i valori riportati sono riferibili a tutte le specie legnose, con scarti del 2-3% di umidità per le specie più ricche di estrattivi.
Umidità relativa dell'aria |
Temperature (°C) |
|||||||||
|
0° |
10° |
20° |
30° |
40° |
50° |
60° |
70° |
80° |
90° |
5 % |
1 % |
1 % |
1 % |
1 % |
1 % |
1 % |
1 % |
1 % |
1 % |
1 % |
10 % |
3 % |
3 % |
3 % |
2 % |
2 % |
2 % |
2 % |
2 % |
1 % |
1 % |
15 % |
4 % |
4 % |
4 % |
3 % |
3 % |
3 % |
3 % |
2 % |
2 % |
2 % |
20 % |
5 % |
5 % |
5 % |
4 % |
4 % |
4 % |
3 % |
3 % |
3 % |
2 % |
25 % |
6 % |
5 % |
5 % |
5 % |
5 % |
5 % |
4 % |
4 % |
3 % |
3 % |
30 % |
6 % |
6 % |
6 % |
6 % |
6 % |
5 % |
5 % |
4 % |
4 % |
3 % |
35 % |
7 % |
7 % |
7 % |
7 % |
6 % |
6 % |
5 % |
5 % |
4 % |
4 % |
40 % |
8 % |
8 % |
8 % |
7 % |
7 % |
7 % |
6 % |
6 % |
5 % |
4 % |
45 % |
9 % |
9 % |
9 % |
8 % |
8 % |
7 % |
7 % |
6 % |
6 % |
5 % |
50 % |
10 % |
10 % |
9 % |
9 % |
9 % |
8 % |
7 % |
7 % |
6 % |
6 % |
55 % |
11 % |
10 % |
10 % |
10 % |
9 % |
9 % |
8 % |
7 % |
7 % |
6 % |
60 % |
12 % |
11 % |
11 % |
11 % |
10 % |
10 % |
9 % |
8 % |
7 % |
7 % |
65 % |
13 % |
12 % |
12 % |
12 % |
11 % |
10 % |
10 % |
9 % |
8 % |
8 % |
70 % |
14 % |
14 % |
13 % |
13 % |
12 % |
11% |
11 % |
10 % |
9 % |
8 % |
75 % |
15 % |
15 % |
15 % |
14 % |
13 % |
13 % |
12 % |
11 % |
10 % |
9 % |
80 % |
17 % |
17 % |
16 % |
16 % |
15 % |
14 % |
14 % |
13 % |
12 % |
11 % |
85 % |
19 % |
19 % |
18 % |
18 % |
17 % |
16 % |
15 % |
14 % |
13 % |
12 % |
90 % |
22 % |
22 % |
21 % |
20 % |
19 % |
18 % |
17 % |
16 % |
15 % |
14 % |
95 % |
27 % |
26 % |
25 % |
24 % |
23 % |
22 % |
21 % |
20 % |
19 % |
18 % |
100 % |
33 % |
32 % |
31 % |
30 % |
29 % |
28 % |
27 % |
26 % |
25 % |
24 % |
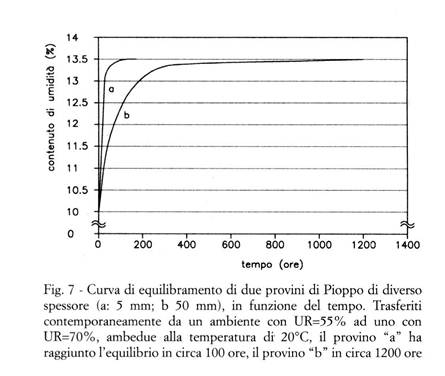
segatura, o comunque particelle nelle quali il rapporto fra superficie e volume è elevato; è invece lungo, talora lunghissimo, in caso di segati o manufatti, nei quali le molecole di acqua devono attraversare un certo spessore di legno per passare dall'atmosfera alle pareti delle cellule, e viceversa ( ). Nella Fig. 7 sono messe a confronto le curve di equilibramento di due provini di Pioppo, trasferiti contemporaneamente da un ambiente con UR=55% ad uno con UR=70%, ambedue alla temperatura di 20°C (come risulta dalla Tab. 2, a tali valori corrispondono umidità di equilibrio del legno, rispettivamente, di circa 10% e 13%): il provino "a", spesso 5 mm, ha raggiunto l'equilibrio (U=13,5%) in circa 100 ore, il provino "b", spesso 50 mm, in circa 1200 ore. A causa della continua ed inevitabile variazione (stagionale, diurna, etc.) delle condizioni termoigrometriche ambientali, è raro che un pezzo di legno si trovi in perfetto equilibrio igroscopico con l’ambiente, ed all'interno dei manufatti lignei si instaurano praticamente sempre dei gradienti di umidità ( ) (v. il § 3.3.5).
Ritiri e rigonfiamenti
Mentre in molti altri materiali (e soprattutto nei metalli) le maggiori variazioni dimensionali sono prodotte dalle variazioni di temperatura, nel legno i coefficienti di dilatazione termica sono assai piccoli, ed in genere trascurabili rispetto alle variazioni dimensionali indotte dalle variazioni di umidità che si verificano nell'intervallo igroscopico.
Tab. 3 - Valori tipici dei ritiri totali (fra stato fresco e stato anidro, espressi come variazione dimensionale percentuale rispetto alla dimensione fresca) nelle direzioni anatomiche trasversali, per alcuni legni (da: Fioravanti, 1994; Hoadley, 1998)
|
|
Ritiro tangenziale |
Ritiro radiale |
(da: Fioravanti, 1994) |
|
|
|
Faggio |
Fagus sylvatica L. |
10 |
5,8 |
Farnia, Rovere |
Quercus sp.p. |
8,3 |
4,4 |
Gattice (= Pioppo bianco) |
Populus alba L. |
6,3 |
2,8 |
Noce |
Juglans regia L. |
7,3 |
5 |
Tiglio |
Tilia cordata Mill. |
9,2 |
5,6 |
Abete bianco |
Abies alba Mill. |
7,2 |
3,5 |
Abete rosso |
Picea abies Karst. |
8,5 |
4 |
Pino silvestre |
Pinus sylvestris L. |
7 |
4 |
(da: Hoadley, 1998) |
|
|
|
Spruce (Abete rosso) |
Picea spp. |
7,4 |
3,6 |
Fir (Abete bianco) |
Abies spp. |
7,6 |
3,8 |
Pine, Scots (Pino silvestre) |
Pinus sylvestris |
7,7 |
4,0 |
Larch (Larice) |
Larix spp. |
7,8 |
3,3 |
Mahogany (Mogano) |
Swietenia spp. |
5,1 |
3,2 |
Walnut, European (Noce) |
Juglans regia |
6,4 |
4,3 |
Chestnut (Castagno) |
Castanea spp. |
6,8 |
4,0 |
Willow (Salice) |
Salix spp. |
7,2 |
4,2 |
Alder (Ontano) |
Alnus spp. |
7,3 |
4,4 |
Cherry (Ciliegio) |
Prunus spp. |
7,8 |
4,2 |
Ash (Frassino) |
Fraxinus spp. |
8,3 |
5,2 |
Poplar (Pioppo) |
Populus spp. |
8,5 |
3,4 |
Maple (Acero) |
Acer spp. |
8,8 |
4,2 |
Elm (Olmo) |
Ulmus spp. |
9,1 |
5,2 |
Lime (Tiglio) |
Tilia spp. |
9,5 |
6,8 |
Oak, White (Quercia bianca) |
Quercus spp. |
10,2 |
5,2 |
Beech (Faggio) |
Fagus spp. |
11,8 |
5,8 |
I ritiri e rigonfiamenti del legno sono fortemente influenzati dalla sua anisotropia, e possono causare sia importanti variazioni dimensionali sia variazioni di forma, tensioni interne, deformazioni permanenti, fessurazioni.
Variazioni dimensionali
Si definisce ritiro dimensionale totale (o ritiro lineare totale) βDtot la diminuzione dimensionale percentuale che un pezzo di legno subisce lungo una generica direzione D quando la sua umidità passa dallo stato fresco (cioè maggiore del punto di saturazione) allo stato anidro:
βDtot = [(Dfr - Do) / Dfr ] x 100
Data la anisotropia del legno, risulta conveniente considerare i ritiri dimensionali totali βLtot , βRtot , βTtot , secondo le tre direzioni anatomiche fondamentali ( ):
- il ritiro longitudinale βLtot (tipicamente compreso fra 0,2% e 0,8%) è molto piccolo, ed in molti casi può essere trascurato ( )
- il ritiro radiale βRtot è tipicamente compreso fra 3% e 6%
- il ritiro tangenziale βTtot è tipicamente compreso fra 6% e 12%
Con discreta approssimazione si può ritenere che i ritiri parziali (cioè fra lo stato fresco ed una umidità finale Ufin diversa da zero) siano proporzionali alla variazione di umidità intercorrente fra il punto di saturazione Usat e l'umidità finale Ufin (si considera Usat perché il ritiro comincia a manifestarsi soltanto a partire da tale umidità).
Considerazioni analoghe possono farsi per il rigonfiamento del legno, che con buona approssimazione (a parte l'isteresi) può essere trattato come un ritiro di segno opposto.
Il passare degli anni e l'esposizione a continue variazioni termoigrometriche ambientali possono talora ridurre leggermente l'igroscopicità del legno ed i suoi valori di ritiro ( ), ma mai eliminarli completamente (a meno che non sopravvengano gravi degradamenti, p.es. quelli prodotti da elevate temperature, o da attacchi di funghi od insetti).
Un semplice calcolo per stimare l'entità dei ritiri
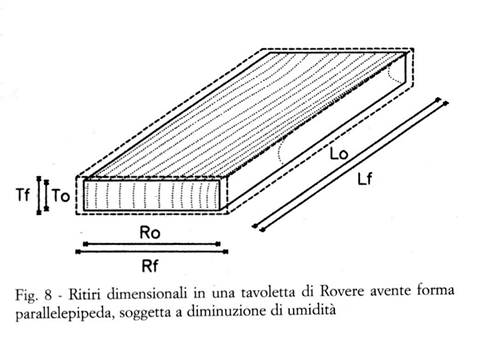
Si consideri ad esempio la tavoletta di Rovere della Fig. 8, avente forma parallelepipeda, con gli spigoli orientati secondo le tre direzioni anatomiche principali, avente i ritiri totali e le dimensioni seguenti:
lunghezza (direzione longitudinale) 800 mm βLtot = 0,1 %
larghezza (direzione radiale) 200 mm βRtot = 5,3 %
spessore (direzione tangenziale) 30 mm βTtot = 10,5 %
Passando dall'umidità iniziale U=90% (stato fresco) all'equilibrio in un ambiente umido (UR=80%; Ueq=16%, v. la Tab. 2) la variazione di umidità che produce ritiri vale
ΔU = (Usat - Ueq) = 30% - 16% = 14%
ed i ritiri causati da tale variazione valgono i 14/30 dei corrispondenti ritiri totali; quindi la tavoletta subisce le seguenti diminuzioni dimensionali:
in lunghezza: i 14/30 dello 0,1% di 800 mm, cioè 800x0,001x14/30 = 0,37 mm
in larghezza: i 14/30 del 5,3% di 200 mm, cioè 200x0,053x14/30 = 4,95 mm
in spessore: i 14/30 del 10,5% di 30 mm, cioè 30x0,105x14/30 = 1,47 mm
Se la stessa tavoletta viene poi portata ad equilibrarsi in un ambiente più asciutto (UR=50%; Ueq=9%, v. la Tab. 2), l'ulteriore variazione di umidità ΔU = 16% - 9% = 7% produce le seguenti ulteriori variazioni dimensionali (cioè variazioni dimensionali rispetto allo stato equilibrato precedente):
in lunghezza: i 7/30 dello 0,1% di 800 mm, cioè 800x0,001x7/30 = 0,19 mm
in larghezza: i 7/30 del 5,3% di 200 mm, cioè 200x0,053x7/30 = 2,47 mm
in spessore: i 7/30 del 10,5% di 30 mm, cioè 30x0,105x7/30 = 0,73 mm
I calcoli sopra riportati costituiscono delle approssimazioni ragionevoli delle variazioni dimensionali di una tavoletta ideale, perfettamente orientata secondo le direzioni anatomiche, presentante valori di ritiro noti ed omogenei (cioè uguali in ogni sua parte), fatta equilibrare lentamente (evitando così l'insorgere di tensioni interne), capace di deformarsi liberamente (v. il § Errore. L'origine riferimento non è stata trovata.). In molti casi reali la stima risulterà forse meno accurata, ma potrà comunque fornire almeno degli ordini di grandezza.
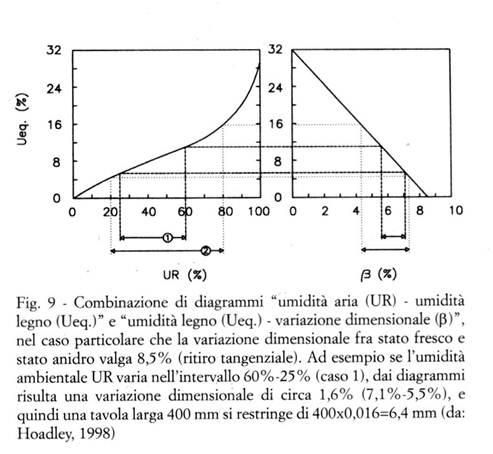
I due diagrammi combinati riportati nella Fig. 9 consentono, in base agli stessi procedimenti, di stimare direttamente le variazioni dimensionali percentuali di elementi lignei non vincolati, quando si equilibrano da una condizione ambientale ad un'altra.
Conseguenze del ritiro, per il legno
Quando l'umidità del legno cambia nell'intervallo igroscopico, a seguito di variazioni termoigrometriche ambientali, il fenomeno dei ritiri è di per sé inevitabile.
Per meglio analizzarne le singole manifestazioni, le conseguenze dei ritiri possono essere schematicamente classificate come "inevitabili" oppure "evitabili" ( ) ( ):
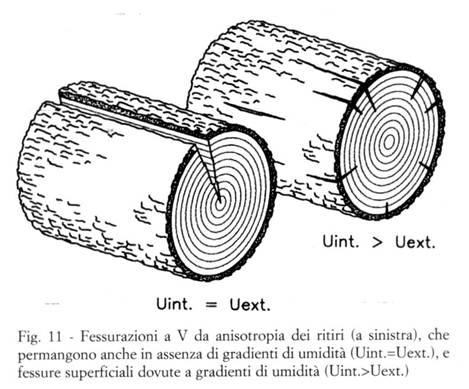
- conseguenze "inevitabili" dovute alla anisotropia del legno: deformazione della sezione trasversale di qualsiasi pezzo di legno, fessure radiali a "V" negli assortimenti contenti il midollo del tronco, imbarcamento delle tavole tangenziali;
- conseguenze "inevitabili" dovute alla presenza di anomalie del legno (legno di reazione, legno giovanile, fibratura elicoidale, etc.): arcuature, falcature, svergolamenti;
- conseguenze "inevitabili" dovute a disomogeneità del legno: p.es. distacchi fra legno primaverile e legno tardivo, causati da differenze di ritiro derivanti da differenze di densità;
- conseguenze "evitabili" dovute a gradienti di umidità: fessurazioni radiali, tensioni interne, cretti sulle testate, collassi.
Talora il legno tende soltanto a deformarsi, in quanto la effettiva deformazione è impedita da vincoli, che possono essere di vario genere: vincoli esterni al pezzo, che ne impediscono la deformazione (p.es. le traverse applicate dietro ai supporti lignei dei dipinti su tavola); vincoli interni, costituiti dalla continuità materiale di uno stesso pezzo o di più pezzi intimamente collegati, le cui diverse parti tendono a deformarsi diversamente l'una dall'altra. In ogni caso, alle deformazioni impedite corrisponde l'insorgere di tensioni interne al materiale; a loro volta tali tensioni possono produrre rotture e/o ulteriori deformazioni.
Deformazioni e fessurazioni causate dall'anisotropia dei ritiri
A causa dell'anisotropia dei ritiri (e dei rigonfiamenti), le variazioni di umidità producono in genere nei manufatti lignei non soltanto variazioni dimensionali, ma anche deformazioni.
Se infatti il ritiro radiale e quello tangenziale fossero uguali, al variare dell'umidità le dimensioni un generico pezzo di legno aumenterebbero o diminuirebbero in misura uguale fra loro, e la sua forma rimarrebbe geometricamente simile a sé stessa; dato che invece il ritiro tangenziale è circa doppio di quello radiale, e quest’ultimo è 10-15 volte maggiore di quello assiale, la forma del pezzo si modifica. In pratica è particolarmente importante la deformazione della sezione trasversale, che si distorce con modalità diverse a seconda dell'orientamento degli anelli di accrescimento nella sezione stessa.
Dalla Fig. 10 risulta ad esempio evidente che nelle tavole tangenziali gli anelli di accrescimento formano angoli diversi con le due facce della tavola, e che in ciascuno di essi la faccia "esterna" (cioè quella più lontana dal midollo del tronco) si avvicina alla direzione tangenziale più della faccia "interna": si può quindi intuire (e si potrebbe dimostrarlo rigorosamente, combinando mediante somme vettoriali le componenti di ritiro radiali e tangenziali) che in sezione trasversale la faccia esterna ritira più di quella interna, e che tale differenza di ritiro fra le due facce provoca l'imbarcamento della tavola (v. la Fig. 10). Al contrario, per evidenti motivi di simmetria, una tavola perfettamente radiale rimane piana e non si imbarca.
Nel caso invece di una rotella (sezione trasversale intera del tronco) od addirittura di un tronco, a causa dell'anisotropia dei ritiri la circonferenza della sezione (orientata in ogni punto secondo la direzione tangenziale) tende ad accorciarsi più del corrispondente raggio (orientato secondo la direzione radiale); essendo impossibile che tali deformazioni risultino compatibili fra loro, nella rotella insorgono delle tensioni di trazione in direzione tangenziale, che portano il legno a deformarsi od addirittura a rompersi.
Questo tipo di fessurazioni si manifesta “inevitabilmente”, a seguito della normale stagionatura (anche se condotta lentamente e con ogni cautela), in qualsiasi pezzo di legno massiccio che contenga il midollo del tronco ( ). La rottura avviene in genere lungo superfici radiali (che proprio a causa dei raggi parenchimatici sono meno resistenti a trazione trasversale), e si manifesta come una o più fessurazioni longitudinali-radiali, che seguono l’andamento della fibratura ed hanno forma di “V” col vertice nel midollo (v. la Fig. 11).
Tipici esempi di queste fessurazioni longitudinali causate dall’anisotropia dei ritiri possono essere riscontrati nelle travi di strutture portanti, o nelle sculture lignee: essendo necessari per questi tipi di manufatto pezzi con dimensione trasversale relativamente grande, viene comunemente usata l’intera sezione del tronco ( ).
Deformazioni e fessurazioni causate da gradienti e differenze di umidità
Se un pezzo di legno si trova in un ambiente le cui condizioni termoigrometriche sono rimaste rigorosamente costanti per lungo tempo, si può ritenere che esso sia completamente equilibrato in relazione a quelle condizioni, e che l'umidità del legno sia uguale in ogni suo punto, indipendentemente dalla maggiore o minore distanza dalla superficie.
Se le condizioni ambientali variano, inizia a ristabilirsi un equilibrio con i nuovi valori. Il pezzo di legno non può portarsi completamente ed immediatamente nelle nuove condizioni di equilibrio (come invece potrebbe avvenire per un truciolo di legno sottilissimo e ben ventilato): per primo giunge alle condizioni di equilibrio lo strato superficiale, poi le variazioni di umidità si propagano più o meno lentamente in profondità. Fino a che il processo di equilibramento non è completato, nello spessore del legno si realizzano inevitabilmente dei gradienti di umidità (differenze di umidità fra strati adiacenti), più o meno elevati.
Se le nuove condizioni ambientali rimangono costanti, dopo un tempo sufficientemente lungo ( ) l'umidità del legno si equalizza in tutto lo spessore, ed i gradienti di umidità si annullano. Se invece le condizioni ambientali variano continuamente, il legno non ha il tempo di raggiungere l'equilibrio igroscopico in tutto lo spessore; anche intuitivamente, senza necessità di ricorrere a complessi modelli matematici, è evidente che gli strati superficiali seguiranno le fluttuazioni ambientali più fedelmente e più rapidamente rispetto agli strati interni, e che i gradienti di umidità avranno andamento complesso e variabile nel tempo.
In conseguenza dei suddetti gradienti di umidità (oppure di differenze di umidità fra pezzi adiacenti), strati o parti di legno adiacenti tendono a ritirarsi o rigonfiarsi di quantità diverse (gradienti di deformazione): ciò provoca nel legno tensioni interne ed ulteriori tendenze a distorcersi.
Dalla sia pur sommaria analisi svolta, risulta evidente che in pratica i gradienti e le differenze di umidità nei manufatti lignei risultano inevitabili, nella misura in cui lo sono le variazioni dell’umidità ambientale.
Comportamento meccanico
Sotto l'azione di sollecitazioni ( ), il legno manifesta deformazioni ed eventualmente anche rotture.
Come molte altre proprietà, anche il comportamento meccanico del legno è fortemente anisotropo, cioè risulta assai diverso a seconda delle direzioni anatomiche secondo le quali vengono applicate le sollecitazioni (Tab. 4).
Nelle applicazioni strutturali è particolarmente importante il comportamento del legno nella direzione longitudinale, secondo la quale risultano massime la sua resistenza e la sua rigidezza; nei supporti lignei dei dipinti su tavola invece assume importanza preponderante il comportamento nelle direzioni trasversali (quella radiale, quella tangenziale, e le infinite direzioni intermedie). In tali direzioni infatti rigidezza e resistenza meccanica risultano minime, mentre è massima la deformabilità causata dalle variazioni di umidità; con le variazioni termoigrometriche ambientali i supporti tendono quindi a deformarsi, rischiando di danneggiare sia la propria forma e la propria integrità, sia la conservazione degli strati pittorici.
Tab. 4 - Valori tipici di alcune proprietà meccaniche in direzione longitudinale e trasversale, determinati su piccoli provini privi di anomalie e difetti (da: Dinwoodie, 1989)
Specie |
Densità |
Umidità |
EL |
ER |
ET |
Balsa (Balsa) |
200 |
9 |
6 274 |
296 |
103 |
Walnut (Noce) |
590 |
11 |
11 239 |
1 172 |
621 |
Ash (Frassino) |
670 |
9 |
15 790 |
1 516 |
827 |
Beech (Faggio) |
750 |
11 |
13 700 |
2 240 |
1 140 |
Sitka spruce (Abete di Sitka) |
390 |
12 |
11 583 |
896 |
496 |
Scots pine (Pino silvestre) |
550 |
10 |
16 272 |
1 103 |
573 |
Douglas fir (Douglasia |
590 |
9 |
16 400 |
1 300 |
900 |
EL Modulo elastico (di Young) in direzione longitudinale (L)
ER Modulo elastico (di Young) in direzione radiale (R)
ET Modulo elastico (di Young) in direzione tangenziale (T)
Specie |
Densità |
Umidità |
RT,LONG |
RT,TRASV |
RC,LONG |
RC,TRASV |
Balsa (Balsa) |
180 |
12 |
35 |
1,6 |
16 |
0,9 |
Ash (Frassino) |
689 |
12 |
136 |
- |
53 |
10,1 |
Beech (Faggio) |
690 |
12 |
180 |
- |
56 |
10,5 |
Sitka spruce (Abete di Sitka) |
433 |
12 |
139 |
2,6 |
39 |
4,1 |
Scots pine (Pino silvestre) |
510 |
12 |
110 |
- |
47 |
5,5 |
Douglas fir (Douglasia) |
545 |
12 |
138 |
2,9 |
50 |
6,9 |
RT,LONG Resistenza a trazione longitudinale
RT,TRASV Resistenza a trazione trasversale
RC,LONG Resistenza a compressione longitudinale
RC,TRASV Resistenza a compressione trasversale
Comportamento elastico, viscoelastico e meccano-sorbitivo
Per una comprensione anche semplicemente intuitiva del comportamento meccanico dei supporti lignei, conviene analizzare separatamente i tre seguenti comportamenti del legno (elastico, viscoelastico, meccanosorbitivo), che complessivamente costituiscono il comportamento reologico; nelle situazioni reali tali comportamenti possono coesistere e manifestarsi congiuntamente, potendo l'uno o l'altro prevalere a seconda delle circostanze.
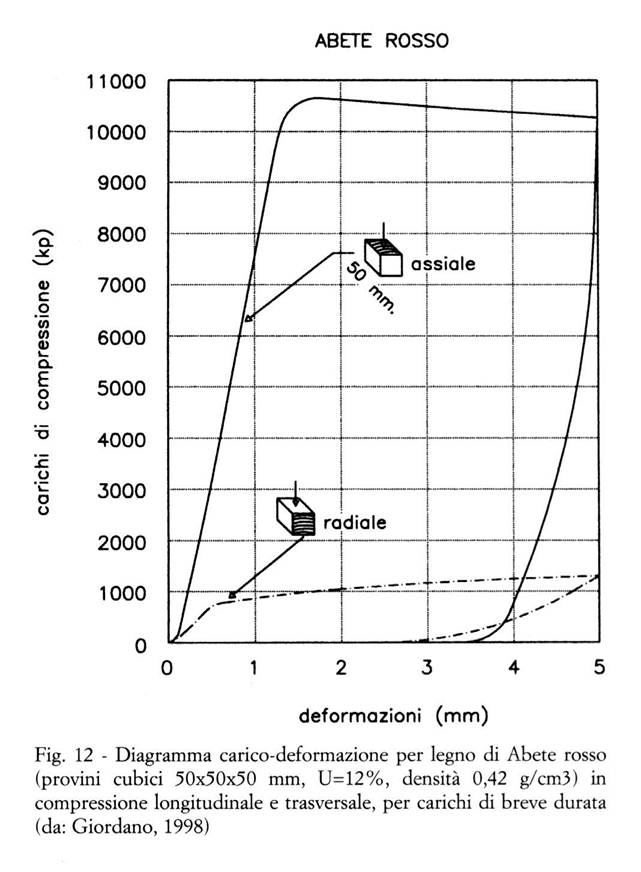
Il comportamento elastico ( ) può essere schematicamente rappresentato su un diagramma carico-deformazione per carichi di breve durata (Fig. 12):
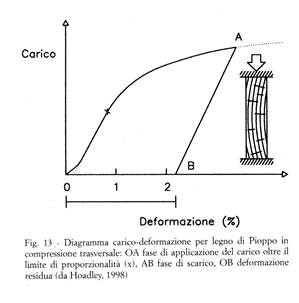
- secondo un andamento ben noto, simile p.es. a quello dei metalli, al crescere del carico applicato la deformazione cresce, dapprima proporzionalmente al carico (fino al limite di proporzionalità), quindi più che proporzionalmente (fino al limite di elasticità), poi subisce delle deformazioni plastiche (irreversibili), infine giunge a rottura;
- se il carico non supera il limite di elasticità, ma viene ridotto fino ad annullarsi, anche la deformazione torna a zero (deformazione residua nulla, che nel legno può avvenire soltanto per carichi di breve durata; per carichi di durata non breve entra inevitabilmente in gioco anche lo scorrimento, v. oltre);
- l'inclinazione del tratto rettilineo iniziale del diagramma (che in tale tratto segue la legge di Hooke) rappresenta la deformabilità del materiale: quanto meno esso è deformabile, tanto più l'inclinazione si avvicina alla verticale, tanto più elevato è il valore numerico del suo modulo elastico E (o modulo di Young);
- se la deformazione plastica prima della rottura è grande, il comportamento a rottura del materiale viene detto duttile; se prima della rottura tale deformazione plastica è molto piccola, od addirittura inesistente, il comportamento a rottura viene invece detto fragile.
Per quanto riguarda il legno, tipicamente:
- in direzione longitudinale presenta modulo elastico e limite elastico elevati, comportamento a flessione duttile per il legno netto (cioè privo di difetti), fragile per il legno con difetti importanti;
- in direzione trasversale presenta modulo elastico e limite elastico bassi, comportamento duttile a compressione (Fig. 13) e fragile a trazione;
- al crescere dell'umidità (nel campo igroscopico) diminuiscono modulo elastico e limite elastico, aumenta la duttilità.
Il comportamento viscoelastico si manifesta quando carichi costanti restano applicati per un tempo più o meno lungo, in condizioni di temperatura ed umidità costanti (se queste variano, subentra il comportamento meccano-sorbitivo, v. oltre), ed è caratterizzato dall'aumento progressivo delle deformazioni col trascorrere del tempo, denominato scorrimento (in inglese: creep) ; esso si manifesta in vari materiali, e nel legno è tanto più accentuato quanto più sono elevate l'umidità, la temperatura, e la tensione applicata. Strettamente correlato ad esso è il rilassamento, caratterizzato dal fatto che imponendo al materiale una deformazione fissa, la corrispondente sollecitazione del materiale progressivamente diminuisce.
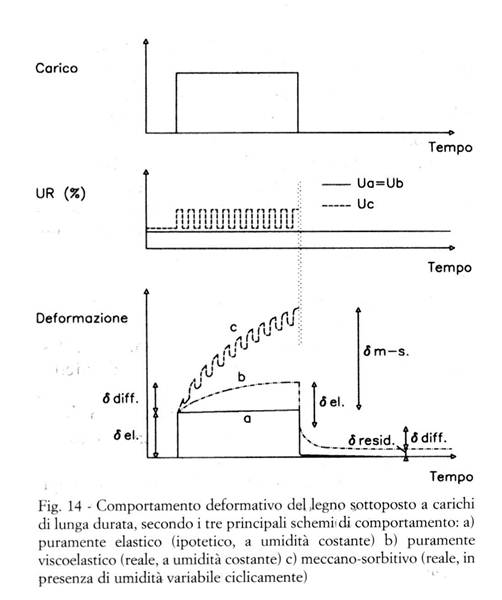
A titolo di esempio, nella Fig. 14 la curva "b" rappresenta l'andamento (viscoelastico) della deformazione, quando il carico viene applicato istantaneamente, rimane costante per un certo periodo, quindi viene istantaneamente azzerato, in condizioni di umidità e temperatura costanti:
- applicando istantaneamente il carico, si ottiene una deformazione istantanea (sostanzialmente corrispondente alla deformazione elastica);
- mantenendo il carico costante, alla deformazione istantanea si aggiunge una deformazione differita, crescente nel tempo (aumentando fino a rottura, oppure crescendo asintoticamente verso un valore limite, a seconda dell'entità del carico in relazione alle caratteristiche del materiale);
- azzerando istantaneamente il carico, la deformazione presenta una brusca diminuzione di tipo elastico (recupero istantaneo), seguita da una diminuzione più lenta (recupero differito), e tende asintoticamente ad una deformazione permanente (anche se il carico non ha mai superato il limite di elasticità, purché abbia agito per tempo sufficientemente lungo; se viceversa il carico ha agito per breve tempo, la deformazione residua vale zero).
Il comportamento meccano-sorbitivo si manifesta quando il legno subisce delle variazioni di umidità e/o temperatura mentre è anche sollecitato meccanicamente; a titolo di esempio, nella curva "c" della Fig. 14 viene rappresentato l'andamento della deformazione quando il carico rimane applicato mentre l'umidità del legno varia ciclicamente:
- quando è sottoposto contemporaneamente a sollecitazioni meccaniche (soprattutto in direzione trasversale) ed igrotermiche, il legno presenta deformazioni significativamente maggiori (e perviene a rottura per carichi minori) che non quando è sottoposto a sollecitazioni puramente meccaniche;
- in tali casi il comportamento è complesso, e tuttora oggetto di ampie ed accurate ricerche (Grill 1996; Svensson e Toratti 1997).
Bibliografia per approfondimenti
GIORDANO G., 1980 - I legnami del mondo (Dizionario enciclopedico). Il Cerilo, Roma
GIORDANO G., 1981- 1988- Tecnologia del Legno. (Vol. 1: La materia prima, 1981 - Vol. 2* e 2**: Le lavorazioni industriali, 1983 - Vol. 3*: I miglioramenti, le prove, l’utilizzazione dei cascami, 1986 - Vol. 3**: I legnami del commercio, 1988) UTET, Torino
NARDI BERTI R., 1979 - La struttura anatomica del legno ed il riconoscimento dei legnami italiani di più corrente impiego. CNR - Istituto del Legno, CSP n. XXIV, Firenze (Ristampe 1982 e 1993)
UNI 2853-1973 - Nomenclatura delle specie legnose che vegetano spontanee in Italia. (Aggiornata con UNI FA 147-84) UNI, Milano
UNI 2854-1987 - Nomenclatura delle specie legnose esotiche coltivate in Italia. UNI, Milano
UNI 3917-1983 - Nomenclatura commerciale dei legnami esotici d’importazione. (Aggiornata con UNI FA 1-89) UNI, Milano
UZIELLI L., 1995 - La conoscenza delle specie legnose - In: “Progetto cultura: il legno nell’Arte (Regione Umbria)”, Federlegno-Arredo e Ministero delle Risorse Agricole Alimentari e Forestali, XXXIX-L
TSOUMIS G. , 1991 – Science and Technologyof Wood – Van Nonstrand Reinhold , New York – ISBN 0-422-23985-8
Note di chiusura
Le condizioni termoigrometriche ambientali sono definite dalla temperatura e dalla umidità relativa dell'aria. Quest'ultima è il rapporto percentuale fra la quantità di vapore effettivamente contenuta in un dato volume d'aria (ad una data temperatura e pressione), e la quantità massima di vapore che potrebbe esservi contenuta (nelle stesse condizioni di temperatura e pressione); essa può quindi variare fra lo 0% ed il 100% (condizioni di saturazione).
Vale la pena di sottolineare che l'umidità relativa dell'aria e l'umidità del legno, per quanto ambedue espresse in percentuale, rappresentano grandezze assolutamente diverse, e non coincidenti numericamente; il raggiungimento dell'equilibrio igroscopico fra aria e legno non significa quindi che umidità del legno ed umidità dell'aria abbiano lo stesso valore.
Fra i molteplici fattori che influenzano la velocità degli scambi di umidità fra legno ed atmosfera circostante possiamo qui citare, senza alcuna pretesa di completezza, la specie legnosa - le dimensioni del pezzo - le direzioni e sezioni anatomiche delle superfici esposte, e la loro finitura superficiale - se si tratti di alburno o di durame - la presenza di vernici, rivestimenti, sporcizia, od altre sostanze - temperatura ed umidità del legno e dell'aria - velocità dell'aria, etc.
Si definisce “gradiente di umidità” il rapporto tra la differenza delle umidità del legno in due zone adiacenti, e la distanza fra tali due zone: a parità di distanza, un gradiente è tanto più elevato (ripido) quanto maggiore è la differenza di umidità.
Valgono quindi le relazioni seguenti:
in direzione longitudinale: βLtot = [(Lfr - Lo) / Lfr ] x 100
in direzione radiale: βRtot = [(Rfr - Ro) / Rfr ] x 100
in direzione tangenziale: βTtot = [(Tfr - To) / Tfr ] x 100
essendo Lfr , Rfr , Tfr le dimensioni allo stato fresco, ed Lo , Ro , To le dimensioni allo stato anidro, nelle direzioni rispettivamente longitudinale, radiale e tangenziale.
Analogamente si definisce il ritiro volumetrico: βVtot = [(Vfr - Vo) / Vfr ] x 100
che con buona approssimazione può essere calcolato come somma dei tre ritiri dimensionali:
βVtot = βLtot + βRtot + βTtot
In casi particolari (p,es, legno giovanile, legno di reazione), il ritiro longitudinale può essere assai maggiore (10 volte quello del legno normale, ed anche di più), e può provocare gravi tensioni interne e deformazioni, a causa delle diverse variazioni dimensionali fra il legno "normale" e l'adiacente legno "anomalo".
Nel legno "vecchio" (cioè lavorato e messo in opera da molti anni, od addirittura secoli) il continuo alternarsi delle condizioni climatiche cui inevitabilmente è stato esposto tende a produrre una certa riduzione non soltanto dell'entità dei ritiri e rigonfiamenti igroscopici, ma anche dell'ampiezza del ciclo di isteresi (cioè della differenza dei valori di equilibrio fra i cicli di adsorbimento e quelli di desorbimento; v. la Fig. 6).
In realtà la distinzione non è sempre così netta, e le conseguenze qui indicate come "evitabili" lo sono solo parzialmente, anche applicando in modo razionale le migliori tecniche di essiccazione o di condizionamento del legno.
Vengono definite le seguenti quattro principali modalità di deformazione dei segati (v. la Fig. 5): imbarcamento, falcatura, arcuatura e svergolamento. Esse possono essere transitorie o permanenti, e talora due o più di esse possono manifestarsi contemporaneamente in uno stesso segato.
Varie possono esserne le cause, isolate od in combinazione fra loro.
In qualche caso la rottura può non manifestarsi, p.es. se l’umidità di equilibrio è elevata, se la specie legnosa presenta elevata resistenza a trazione trasversale od è particolarmente stabile (rapporto βT / βR piccolo), se il legno si è dilatato (elasticamente o con deformazione permanente) in modo da lasciare almeno in parte rilassare le tensioni; talvolta le fessurazioni rimangono allo stato latente, e si manifestano poi d’improvviso a seguito di ulteriori abbassamenti dell’umidità, vibrazioni, urti, etc.
All’aumentare od al diminuire dell’umidità del legno, queste fessurazioni tendono rispettivamente a restringersi o ad allargarsi. È quindi sconsigliabile cercare di chiuderle o stuccarle con materiali rigidi (legno, stucco rigido, resina, etc.), perché se la tendenza della fessura a restringersi risulta impedita, ne derivano tensioni che possono far estendere la fessura stessa in zone di legno ancora integre; se invece la fessura tende ad allargarsi, potranno prodursi crepe fra legno e riempimento, oppure (se il riempimento è robusto e ben aderente) rotture in altre parti del legno. Qualora sia necessario chiudere o mascherare tali fessurazioni (p.es. per motivi estetici), converrà quindi farlo con materiale cedevole, che possa accompagnare le inevitabili deformazioni del legno.
Gli scambi igroscopici fra ambiente e legno possono essere più o meno rapidi, e dipendono da vari fattori (specie legnosa; sezione anatomica apparente sulla superficie di scambio; ventilazione, temperatura, forma, finitura della superficie di scambio; presenza di eventuali strati superficiali che possano fungere da "barriera" all'umidità, quali strato pittorico, cera, vernice, "patina" accumulatasi nel tempo, etc.). Anche i movimenti dell'umidità all'interno del legno - e di conseguenza i relativi gradienti - dipendono da numerosi fattori, fra cui specie legnosa, presenza di alburno o di durame, direzione anatomica, temperatura, presenza di estrattivi o di impregnanti, etc.
Le sollecitazioni possono essere prodotte non soltanto da azioni applicate dall'esterno, ma anche da reazioni vincolari che tendono a contrastare deformazioni (p.es. causate da variazioni di umidità) non consentite da vincoli esterni (p.es. una traversatura, che impedisce l'imbarcamento di un tavolato), oppure interni (p.es. dilatazioni tendenzialmente diverse in zone di legno adiacenti che si trovano ad umidità diverse: tali tensioni interne, in elementi lignei soggetti a gradienti di umidità, sono determinate dalla continuità stessa del materiale).
Il termine "elasticità" viene spesso usato in modo ambiguo e fuorviante, tanto che di frequente si sente dire (a torto) che un materiale è tanto più "elastico" quanto più è cedevole alle sollecitazioni applicate. E' invece opportuno, almeno in ambito tecnico, utilizzare il termine "elasticità" soltanto per indicare la proprietà di un corpo di tornare immediatamente alle dimensioni iniziali una volta cessate le sollecitazioni che lo avevano deformato, e quindi come contrario di "plasticità" (che indica invece la capacità di mantenere permanentemente le deformazioni, anche dopo che sono cessate le sollecitazioni).
Il termine "elastico" viene talora usato per rappresentare il comportamento di un materiale che sopporta elevate deformazioni senza rompersi né deformarsi permanentemente (esempio: il legno di Frassino "è molto elastico"); anche in questo caso si tratta di un uso improprio, perché tale comportamento va invece descritto come "ad elevato limite elastico".
Fonte: http://ottimari.agr.unifi.it/~fioravanti/Tecnologia%20del%20legno%20AA%2009%2010/dispense%20xilo.doc
Autore : Prof Marco Fioravanti
- Università di Firenze - DISTAF (Dipartimento di Scienze e Tecnologie Ambientali Forestali)
(Rielaborato da : L.Uzielli – M.Fioravanti Il Comportamento fisico meccanico dei supporti lignei dei dipinti su tavola Disegni eseguiti da Lorella Bevilacqua)
Comportamenti fisico meccanici del legno
Visita la nostra pagina principale
Comportamenti fisico meccanici del legno
Termini d' uso e privacy