Appunti di Zoologia
Prof. Biagio D’Aniello
La percezione dei suoni nel mondo animale |
La percezione sensoriale dei fenomeni fisici è generalmente una materia piuttosto ostica. L’argomento, per essere compiutamente compreso, richiede una conoscenza approfondita delle diverse manifestazioni fisiche; le quali, peraltro, sono accompagnate da un formulario matematico alquanto complesso. D’altra parte diventa ancora più complicato se si considera che la sensibilità biologica è un fenomeno strettamente personale e, di conseguenza, ha una componente soggettiva difficile da esprimere in maniera univoca. Ad esempio, la rilevazione di un colore è sicuramente simile per persone non daltoniche, tuttavia, le sue tonalità possono essere percepite in maniera lievemente diversa da due individui. Un analogo discorso vale per la percezione del suono o per alcune sostanze chimiche nel gusto. A tutto ciò si aggiunge che alcuni segnali non possono essere affatto riconosciuti dal nostro sistema sensoriale ed è quantomeno arduo interpretare o immaginare come essi siano avvertiti dalle altre specie animali. Si pensi a tale proposito alla visione degli ultravioletti da parte delle api o alla percezione degli ultrasuoni ad opera dei pipistrelli. Al fine di agevolare lo studio dei meccanismi di comunicazione acustica sono di seguito riportati nelle linee più essenziali le principali caratteristiche del suono e suoi meccanismi di diffusione. |
Cenni sulla natura del suono |
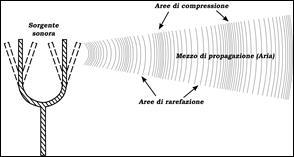 Il suono è fisicamente un fenomeno ondulatorio in un mezzo elastico; pertanto, si tratta di un’energia meccanica. Per la sua realizzazione sono necessarie una sorgente (es. un corpo vibrante) e un mezzo in cui propagarsi (es. aria o acqua). Il fenomeno comporta lo spostamento delle particelle che costituiscono il mezzo di propagazione intorno al punto della loro posizione iniziale (Fig. 1); quindi, la trasmissione del suono è una serie di fenomeni di compressione e decompressione (o rarefazione) che si susseguono della materia in cui si propaga. Il suono è fisicamente un fenomeno ondulatorio in un mezzo elastico; pertanto, si tratta di un’energia meccanica. Per la sua realizzazione sono necessarie una sorgente (es. un corpo vibrante) e un mezzo in cui propagarsi (es. aria o acqua). Il fenomeno comporta lo spostamento delle particelle che costituiscono il mezzo di propagazione intorno al punto della loro posizione iniziale (Fig. 1); quindi, la trasmissione del suono è una serie di fenomeni di compressione e decompressione (o rarefazione) che si susseguono della materia in cui si propaga.
Per come definito il concetto di suono comprenderebbe tutte le manifestazioni meccaniche che si propagano mediante onde; tuttavia, noi generalmente parliamo di suono riferendoci a qualsiasi forma di energia meccanica percepibile dal nostro orecchio. Le nostre capacità percettive rientrano in un determinato ambito di frequenze (vedi dopo), al di sopra delle quali parleremo di ultrasuoni e al di sotto di infrasuoni.
Un rumore si può intendere come una manifestazione sonora fastidiosa o indesiderata, tuttavia, si tratta di un concetto piuttosto soggettivo; infatti, alcuni soggetti percepiscono sgradevolmente certi tipi di musica. Inoltre, anche un suono melodioso ad elevato volume può diventare sgradevole.
Un qualsiasi suono presenta delle proprietà fisiche ben definite che lo caratterizzano.
|
Ampiezza |
 L’ampiezza rappresenta l’entità dello spostamento delle molecole in seguito ad un fenomeno acustico ed e relativa all’intensità del fenomeno (ciò che noi percepiamo come volume). In effetti, all’aumentare dell’ampiezza di un onda sonora, aumenta anche la sua energia e, di conseguenza, la forza con cui colpisce la membrana timpanica. Graficamente l’ampiezza è rappresentata dall’altezza dell’onda misurata sull’asse delle ordinate (Fig. 2). L’intensità si può misurare con le unità di misura dell’energia (es. watt/m2). Tuttavia, per finalità pratiche è di uso più comune il Bel o il decimo di Bel (decibel = dB). L’ampiezza rappresenta l’entità dello spostamento delle molecole in seguito ad un fenomeno acustico ed e relativa all’intensità del fenomeno (ciò che noi percepiamo come volume). In effetti, all’aumentare dell’ampiezza di un onda sonora, aumenta anche la sua energia e, di conseguenza, la forza con cui colpisce la membrana timpanica. Graficamente l’ampiezza è rappresentata dall’altezza dell’onda misurata sull’asse delle ordinate (Fig. 2). L’intensità si può misurare con le unità di misura dell’energia (es. watt/m2). Tuttavia, per finalità pratiche è di uso più comune il Bel o il decimo di Bel (decibel = dB).
|
Altezza |
 L’altezza di un suono è un termine che non si riferisce al suo volume, ma a quanto esso è acuto, e dipende dalla sua frequenza. Quest’ultima rappresenta il numero di oscillazioni che si producono nell’unità di tempo. Si misura in Hertz (Hz), che esprime il numero di vibrazioni che avvengono in un secondo (Fig. 3). Suoni molto acuti hanno una frequenza elevata e quindi sono anche alti; laddove suoni bassi sono caratterizzati da una minore frequenza. Il campo delle frequenze udibili non è illimitato, ma caratteristico di ogni singola specie. L’altezza di un suono è un termine che non si riferisce al suo volume, ma a quanto esso è acuto, e dipende dalla sua frequenza. Quest’ultima rappresenta il numero di oscillazioni che si producono nell’unità di tempo. Si misura in Hertz (Hz), che esprime il numero di vibrazioni che avvengono in un secondo (Fig. 3). Suoni molto acuti hanno una frequenza elevata e quindi sono anche alti; laddove suoni bassi sono caratterizzati da una minore frequenza. Il campo delle frequenze udibili non è illimitato, ma caratteristico di ogni singola specie.
|
Timbro |
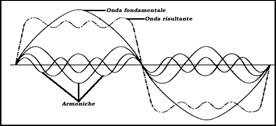 Un qualsiasi suono è quasi sempre accompagnato da una serie di suoni secondari, che negli strumenti musicali sono detti suoni armonici (Fig. 4). Essi si assommano al suono principale deformando l’onda principale. Questo rende l’onda irregolare, ma nello stesso tempo caratteristica. E’ questo il motivo per cui si possono riconoscere i diversi strumenti musicali o le voci anche se hanno la stessa frequenza e intensità. Solo il diapason emette un suono puro, cioè un suono non accompagnato da armoniche. Un qualsiasi suono è quasi sempre accompagnato da una serie di suoni secondari, che negli strumenti musicali sono detti suoni armonici (Fig. 4). Essi si assommano al suono principale deformando l’onda principale. Questo rende l’onda irregolare, ma nello stesso tempo caratteristica. E’ questo il motivo per cui si possono riconoscere i diversi strumenti musicali o le voci anche se hanno la stessa frequenza e intensità. Solo il diapason emette un suono puro, cioè un suono non accompagnato da armoniche.
|
Velocità di propagazione |

Modulo di elasticità e densità possono a loro volta variare in uno stesso mezzo in funzione di altri parametri fisici, come la temperatura (Tabella 2) e, nel caso dell’acqua, la pressione e la salinità. Queste variabili influiscono contemporaneamente sia sul modulo di elasticità, sia sulla densità, in maniera tale che se aumenta la temperatura, la pressione e la salinità di un mezzo di propagazione si accresce anche la velocità del suono.
Diversamente le caratteristiche fisiche del suono non hanno alcun effetto sulla sua velocità di propagazione: un suon molto intenso e alto e uno a scarsa energia e a bassa frequenza avranno la stessa velocità. Ciò che invece cambia al cambiare delle caratteristiche del suono è la distanza di propagazione.
|
Distanza di propagazione |
 La propagazione del suono è limitata da diversi fattori. C’è innanzitutto una dissipazione dell’energia meccanica che si trasforma in calore man mano che l’onda sonora si diffonde a causa dell’attrito con le molecole del mezzo di trasmissione. Inoltre, l’energia del suono allontanandosi dalla sorgente si distribuisce su una superficie sempre più ampia, per cui diviene sempre più rarefatta. S’immagini un emissione sonora da una sorgente puntiforme che può diffondersi senza ostacoli in tutte le direzioni; in questo caso la quantità di energia si distribuisce su una superficie sferica progressivamente maggiore e, pertanto, diviene sempre più rarefatta. Per questo motivo l’emissione del suono in un tubo chiuso, che costringe il suono in una certa direzione, arriva molto più lontano. Allontanandosi dall’emissione si avverte un suono con un’intensità sempre più piccola fino a zero (Fig. 5). La propagazione del suono è limitata da diversi fattori. C’è innanzitutto una dissipazione dell’energia meccanica che si trasforma in calore man mano che l’onda sonora si diffonde a causa dell’attrito con le molecole del mezzo di trasmissione. Inoltre, l’energia del suono allontanandosi dalla sorgente si distribuisce su una superficie sempre più ampia, per cui diviene sempre più rarefatta. S’immagini un emissione sonora da una sorgente puntiforme che può diffondersi senza ostacoli in tutte le direzioni; in questo caso la quantità di energia si distribuisce su una superficie sferica progressivamente maggiore e, pertanto, diviene sempre più rarefatta. Per questo motivo l’emissione del suono in un tubo chiuso, che costringe il suono in una certa direzione, arriva molto più lontano. Allontanandosi dall’emissione si avverte un suono con un’intensità sempre più piccola fino a zero (Fig. 5).
A condizionare la diffusione dei suoni in maniera determinante ci sono molti altri fattori che, talvolta, risultano particolarmente incisivi. Ad esempio il vento, le irregolarità dell’ambiente, gli ostacoli. Il vento non trasporta il suono, ma come gli altri fattori modifica il percorso delle onde. In una stanza la voce si percepisce meglio che all’aperto perché le pareti creano un ostacolo alla diffusione del suono, riflettendolo in una certa misura; in questo modo l’energia tende a rimanere maggiormente presso l’ascoltatore.
 L’effetto di diminuzione varia anche a seconda delle sostanze attraversate. In generale quelle più compatte lo conducono per distanze maggiori. Così nei solidi un suono si propaga per uno spazio maggiore rispetto ad un liquido e ancora di più rispetto ad un gas. Questo è il motivo per cui il rumore di un treno può essere avvertito a maggiore distanza ponendo l’orecchio sulle rotaie. Naturalmente un suono emesso con forte intensità si propagherà di più rispetto ad uno con minore energia (Fig. 6) e lo stesso accade per suoni a bassa frequenza, perché subiscono un minore assorbimento nel mezzo di propagazione (Fig. 7). L’effetto di diminuzione varia anche a seconda delle sostanze attraversate. In generale quelle più compatte lo conducono per distanze maggiori. Così nei solidi un suono si propaga per uno spazio maggiore rispetto ad un liquido e ancora di più rispetto ad un gas. Questo è il motivo per cui il rumore di un treno può essere avvertito a maggiore distanza ponendo l’orecchio sulle rotaie. Naturalmente un suono emesso con forte intensità si propagherà di più rispetto ad uno con minore energia (Fig. 6) e lo stesso accade per suoni a bassa frequenza, perché subiscono un minore assorbimento nel mezzo di propagazione (Fig. 7).
|
In mare l’attenuazione del suono è proporzionale al quadrato della frequenza, quindi gli ultrasuoni si propagano molto di meno rispetto ai suoni a bassa frequenza. Un’interessante fenomeno si verifica nel mare dove la temperatura e la pressione modificano le caratteristiche dell’acqua secondo un gradiente batimetrico, cosicché la velocità del suono è variabile a diverse profondità. Combinando l’effetto della pressione e della temperatura s’individua un punto in profondità in cui la velocità del suono risulta più bassa. Se le onde sonore sono emesse in prossimità del punto di minima velocità si muoveranno naturalmente verso tutte le direzioni. Quelle dirette verso il basso e verso l’alto saranno deviate per un fenomeno di rifrazione verso la linea di minima velocità per il semplice fatto di attraversare materia con uno stato fisico diverso. Così le onde dirette verso l’alto saranno deviate verso il basso e viceversa, sino ad attraversare la linea di minima velocità ed essere nuovamente piegate verso la linea di minima velocità. In sostanza, si crea un canale del suono detto SOFAR (SOund Fixing And Ranging) in cui le onde sono canalizzate anziché dirigersi in tutte le direzioni (Fig. 8). |
Ciò determina una minore dispersione dell’energia e la possibilità per il suono di |
 raggiungere elevatissime distanze. Il fenomeno è paragonabile all’emissione di suoni in un tubo. Il canale SOFAR essendo una funzione della temperatura ed essendo la temperatura variabile con la latitudine e le stagioni, può variare la sua profondità nello spazio (Fig. 9) e nel tempo. Il canale risulta molto più superficiale in mari freddi perché la temperatura del mare è più o meno la stessa lungo la colonna d’acqua, cosicché a determinare le variazioni di velocità del suono è solo la pressione batimentrica, che ne aumenta progressivamente la sua velocità a partire dalla superficie. Il canale SOFAR sembra essere piuttosto importante per la comunicazione dei cetacei a grandi distanze. raggiungere elevatissime distanze. Il fenomeno è paragonabile all’emissione di suoni in un tubo. Il canale SOFAR essendo una funzione della temperatura ed essendo la temperatura variabile con la latitudine e le stagioni, può variare la sua profondità nello spazio (Fig. 9) e nel tempo. Il canale risulta molto più superficiale in mari freddi perché la temperatura del mare è più o meno la stessa lungo la colonna d’acqua, cosicché a determinare le variazioni di velocità del suono è solo la pressione batimentrica, che ne aumenta progressivamente la sua velocità a partire dalla superficie. Il canale SOFAR sembra essere piuttosto importante per la comunicazione dei cetacei a grandi distanze.
|
I suoni nella comunicazione animale |
 La comunicazione serve per modificare il comportamento di un individuo ricevente. Ad esempio un segnale d’allarme induce in un conspecifico una postura d’attenzione. I segnali comunicativi possono essere diretti anche a specie diverse; si pensi all’atteggiamento terrifico assunto da molte specie di mammiferi contro i predatori o competitori, che si ottiene drizzando il pelo dorsale e inarcando il dorso; sono senz’altro più familiari gli atteggiamenti di richiesta del cibo messi in atto dagli animali domestici al fine di condizionare il nostro comportamento a loro favore; oppure l’inequivocabile messaggio contenuto nel ruggito del leone (Fig. 10). La comunicazione serve per modificare il comportamento di un individuo ricevente. Ad esempio un segnale d’allarme induce in un conspecifico una postura d’attenzione. I segnali comunicativi possono essere diretti anche a specie diverse; si pensi all’atteggiamento terrifico assunto da molte specie di mammiferi contro i predatori o competitori, che si ottiene drizzando il pelo dorsale e inarcando il dorso; sono senz’altro più familiari gli atteggiamenti di richiesta del cibo messi in atto dagli animali domestici al fine di condizionare il nostro comportamento a loro favore; oppure l’inequivocabile messaggio contenuto nel ruggito del leone (Fig. 10).
|
Gli animali utilizzano una vasta gamma di segnali per comunicare tra loro. Tra essi la comunicazione acustica ha il vantaggio di raggiungere grandi distanze ed è tanto più utile quando più il campo visivo è limitato. Così in alcuni tipi d’ambiente la comunicazione ottica ha scarsissimo significato. In effetti, in un bosco il fogliame degli alberi e delle piante del sottobosco rendono molto arduo localizzare a distanza altri organismi viventi; analoga situazione si verifica nel mare, in cui la radiazione luminosa è rapidamente assorbita, mentre il suono si può propagare per distanze anche molto superiori al mezzo aereo.
Naturalmente un’emissione acustica diventa un mezzo di comunicazione solo se viene emessa in determinate circostanze e, recepita da un altro animale, ne induce una risposta comportamentale. |
Produzione dei suoni |
 Nonostante la potenziale utilità e l’efficacia dei segnali acustici, la comunicazione sonora è un evento molto sporadico nel regno animale, essendo praticamente limitata agli insetti e ai vertebrati. Tra i vertebrati alcuni pesci sono in grado di produrre suoni; essi sono percepiti dall’orecchio umano come ronzii, soffi, gracidii, grugniti (Fig. 11). La comunicazione sonora è, invece, molto diffusa tra gli anfibi anuri. Ben noto è il gracidio delle rane che può raggiungere distanze considerevoli perché emesso con notevole intensità. Nei rettili la comunicazione sonora è nota in molti gruppi, tra cui i gechi sono sicuramente quelli che presentano una propensione più spiccata verso l’emissione dei suoni (Fig. 12). Tuttavia, seppur più raramente anche i loricati producono suoni, sotto forma di ruggiti durante l’attacco alla preda e i cheloni possono emettere brevi fischi durante l’accoppiamento. Gli uccelli sono ben noti per la loro varietà di canti e versi, peraltro spesso con dialetti locali. Essi sono anche degli ottimi imitatori, come le cince che possono riprodurre in parte i versi delle altre specie; o i pappagalli e i merli indiani che imparano a riprodurre anche alcune parole del linguaggio umano. Tra i mammiferi quasi tutti producono suoni frequentemente e tra essi i più loquaci sono certamente i carnivori e i primati. Essi ne emettono una varietà enorme ognuno dei quali ha un suo significato nella comunicazione. Così si possono sentire bramiti (es. cervidi), ruggiti (es. felidi), nitriti (es. equidi), squittii (es. roditori), ululati (es. canidi), fischi (es. cetacei). Nonostante la potenziale utilità e l’efficacia dei segnali acustici, la comunicazione sonora è un evento molto sporadico nel regno animale, essendo praticamente limitata agli insetti e ai vertebrati. Tra i vertebrati alcuni pesci sono in grado di produrre suoni; essi sono percepiti dall’orecchio umano come ronzii, soffi, gracidii, grugniti (Fig. 11). La comunicazione sonora è, invece, molto diffusa tra gli anfibi anuri. Ben noto è il gracidio delle rane che può raggiungere distanze considerevoli perché emesso con notevole intensità. Nei rettili la comunicazione sonora è nota in molti gruppi, tra cui i gechi sono sicuramente quelli che presentano una propensione più spiccata verso l’emissione dei suoni (Fig. 12). Tuttavia, seppur più raramente anche i loricati producono suoni, sotto forma di ruggiti durante l’attacco alla preda e i cheloni possono emettere brevi fischi durante l’accoppiamento. Gli uccelli sono ben noti per la loro varietà di canti e versi, peraltro spesso con dialetti locali. Essi sono anche degli ottimi imitatori, come le cince che possono riprodurre in parte i versi delle altre specie; o i pappagalli e i merli indiani che imparano a riprodurre anche alcune parole del linguaggio umano. Tra i mammiferi quasi tutti producono suoni frequentemente e tra essi i più loquaci sono certamente i carnivori e i primati. Essi ne emettono una varietà enorme ognuno dei quali ha un suo significato nella comunicazione. Così si possono sentire bramiti (es. cervidi), ruggiti (es. felidi), nitriti (es. equidi), squittii (es. roditori), ululati (es. canidi), fischi (es. cetacei).
|
 Tra gli invertebrati gli ortotteri e gli emitteri sono ben noti per il frinire estivo dei grilli (Fig 14) e delle cicale. Questi suoni si possono udire a notevoli distanze, perché sono spesso presenti sistemi di amplificazione strutturali. In alcuni casi i grilli scavano tane in modo da dargli forme che amplificano il suono. Tra gli invertebrati gli ortotteri e gli emitteri sono ben noti per il frinire estivo dei grilli (Fig 14) e delle cicale. Questi suoni si possono udire a notevoli distanze, perché sono spesso presenti sistemi di amplificazione strutturali. In alcuni casi i grilli scavano tane in modo da dargli forme che amplificano il suono.
|
C’è da dire che molti suoni sono al di fuori della soglia dell’udibile umano che nelle migliori condizioni va dai 20 ai 20.000 Hz. E’ il caso di molti cetacei o pipistrelli le cui emissioni sonore possono essere registrate solo mediante una strumentazione particolare (es. bat detector) e ascoltati dopo essere stati trasformati. In seguito all’analisi degli ultrasuoni degli animali sono anche stati costruiti degli apparecchi che dovrebbero emettere ultrasuoni in grado di disturbare gli animali indesiderati. Tuttavia, essi hanno al momento scarso successo per le notevoli capacità adattative degli animali bersaglio, che possono abituarsi ai suoni. |
L’intensità e la frequenze dei suoni emessi dai cetacei sono molto variabili anche all’interno della stessa specie. I cetacei e i pipistrelli sono in grado di emettere dei suoni con frequenze altissime nel meccanismo di ecolocalizzazione. Sono stati registrate frequenze fino a 150.000 Hz in alcuni cetacei e insetti e leggermente inferiori nei pipistrelli. Comunicazioni nell’ambito dell’ultrasonico sono anche noti in ratti, gerbilli e criceti, ma il significato di questi segnali non è del tutto chiaro. Anche i cani e i gatti sono in grado di percepire gli ultrasuoni, probabilmente perché anche le loro prede comunicano in questo modo e, quindi possono localizzarle. Alcuni esperimenti hanno evidenziato che il gatto risponde a suoni fino a 60.000 Hz ed è noto che molti roditori emettono suoni con frequenza fino a 50.000 Hz. Il cane arriva a percepire frequenze di circa 35.000 Hz. |
 A parte il meccanismo di ecolocalizzazione che richiede frequenze elevate, i cetacei nella normale comunicazione emettono suoni con frequenze più basse: gli odontoceti comunemente usano frequenze intorno ai 25.000 Hz, mentre i misticeti 5.000 Hz. Tuttavia, per le comunicazioni a lunga distanza i cetacei più grandi possono anche usare infrasuoni nell’ordine di poche decine di Hz. I suoni a bassa frequenza sono meno assorbiti e viaggiano a lungo; inoltre, la trasmissione a distanza è favorita anche dalla notevole potenza con cui sono emessi: nel capodoglio (Fig. 15) e nella balenottera comune sono state registrate punte di quasi 200 dB. Se poi i suoni sono emessi nel canale SOFAR, allora essi arrivano a centinaia di chilometri di distanza. Comunicazioni a bassa frequenza sono anche comuni negli elefanti; essi emettono suoni al di sotto dei 20 Hz, perciò non udibili per l’uomo, che si diffondono per alcuni chilometri. I piccioni sono in grado di percepire infrasuoni fino a 0,05 hz. A parte il meccanismo di ecolocalizzazione che richiede frequenze elevate, i cetacei nella normale comunicazione emettono suoni con frequenze più basse: gli odontoceti comunemente usano frequenze intorno ai 25.000 Hz, mentre i misticeti 5.000 Hz. Tuttavia, per le comunicazioni a lunga distanza i cetacei più grandi possono anche usare infrasuoni nell’ordine di poche decine di Hz. I suoni a bassa frequenza sono meno assorbiti e viaggiano a lungo; inoltre, la trasmissione a distanza è favorita anche dalla notevole potenza con cui sono emessi: nel capodoglio (Fig. 15) e nella balenottera comune sono state registrate punte di quasi 200 dB. Se poi i suoni sono emessi nel canale SOFAR, allora essi arrivano a centinaia di chilometri di distanza. Comunicazioni a bassa frequenza sono anche comuni negli elefanti; essi emettono suoni al di sotto dei 20 Hz, perciò non udibili per l’uomo, che si diffondono per alcuni chilometri. I piccioni sono in grado di percepire infrasuoni fino a 0,05 hz.
|
Significato della comunicazione sonora |
L’emissione dei suoni può avere diversi significati. In molti casi il messaggio è stato ampiamente studiato e decifrato dai ricercatori. Noi stessi spesso usiamo un codice sonoro per comunicare con i nostri animali domestici per indurre determinati comportamenti. Così, un fischio può essere un richiamo di ritorno per il proprio cane, o un urlo un incitamento per il cavallo. Ciononostante il significato di alcune emissioni sonore resta ancora un mistero. Basti pensare a questo proposito alla enorme varietà di suoni emessi dai cetacei di cui solo in parte ne conosciamo il senso.
I segnali sonori possono avere diverse funzioni. Di seguito sono riportati alcuni esempi che comprendono le principali vie comunicative del suono. |
Segnali di minaccia e Territorialità |
I segnali di minaccia sono emessi con l’intento di scongiurare un pericoloso scontro fisico, che potrebbe determinare seri danni fisici. La potenza dell’emissione sonora di regola è proporzionale alla stazza dell’animale e al suo stato emotivo. Quindi, di solito uno dei due contendenti rinuncia alla lotta ritirandosi dopo aver valutato l’avversario più forte, o più determinato e aggressivo. La situazione è quella che si crea quando nella nostra specie sono emesse delle urla in una controversia. |
 Molti segnali sonori sono volti a comunicare la proprietà di una certa area, senza arrivare necessariamente all’individuazione visiva dell’eventuale intruso. Difendere un determinato territorio è molto vantaggioso, perché assicura una fonte di cibo da non dividere con altri e talvolta, anche il controllo delle femmine che lo abitano. Il segnale può essere diretto ad un conspecifico, oppure a membri di specie diverse. Spesso la manifestazione acustica è accompagnata da altri messaggi che rafforzano l’efficacia del gesto. Si pensi ad un elefante inferocito che mentre emette il suo barrito carica l’avversario agitando le orecchie e la proboscide (Fig. 16). Si tratta di un segnale inequivocabilmente comprensibile da qualsiasi specie. Esempi di segnali di minaccia nel mondo animale se ne ritrovano in moltissime altre specie: il gatto maschio emette delle urla lamentose durante il periodo della riproduzione per tentare di tenere alla larga i suoi rivali e monopolizzare la femmina. Le scimmie urlatrici in Amazzonia, vari lemuri del Madagascar, i gibboni in Asia delimitano il territorio emettendo grida praticamente per l’intera giornata, ma con maggiore continuità al mattino e alla sera. In verità, i cori dei gibboni danno più la sensazione di suoni melodiosi, che grida scoordinate. I pipistrelli oltre agli ultrasuoni emettono in volo dei segnali udibili per l’uomo che somigliano a degli squittii. Tali vocalizzazioni hanno l’intento di avvertire che quello è il suo territorio di caccia e che non desidera conspecifici estranei. La maggior parte dei canti emessi dagli uccelli servono a segnalare il possesso del territorio. I maschi di alcune falene possono emettere suoni con strutture microscopiche presenti nelle ali, e lo fanno durante la lotta per conquistarsi un luogo adatto per attrarre le femmine. Lo stesso vale per la stridulazione dei grilli e delle cicale che è usata per avvertire altri maschi del possesso del territorio. I grilli maschi sono molto aggressivi tra loro, infatti per allevarli e sentirli cantare è bene mantenerli in gabbie separate. Molti segnali sonori sono volti a comunicare la proprietà di una certa area, senza arrivare necessariamente all’individuazione visiva dell’eventuale intruso. Difendere un determinato territorio è molto vantaggioso, perché assicura una fonte di cibo da non dividere con altri e talvolta, anche il controllo delle femmine che lo abitano. Il segnale può essere diretto ad un conspecifico, oppure a membri di specie diverse. Spesso la manifestazione acustica è accompagnata da altri messaggi che rafforzano l’efficacia del gesto. Si pensi ad un elefante inferocito che mentre emette il suo barrito carica l’avversario agitando le orecchie e la proboscide (Fig. 16). Si tratta di un segnale inequivocabilmente comprensibile da qualsiasi specie. Esempi di segnali di minaccia nel mondo animale se ne ritrovano in moltissime altre specie: il gatto maschio emette delle urla lamentose durante il periodo della riproduzione per tentare di tenere alla larga i suoi rivali e monopolizzare la femmina. Le scimmie urlatrici in Amazzonia, vari lemuri del Madagascar, i gibboni in Asia delimitano il territorio emettendo grida praticamente per l’intera giornata, ma con maggiore continuità al mattino e alla sera. In verità, i cori dei gibboni danno più la sensazione di suoni melodiosi, che grida scoordinate. I pipistrelli oltre agli ultrasuoni emettono in volo dei segnali udibili per l’uomo che somigliano a degli squittii. Tali vocalizzazioni hanno l’intento di avvertire che quello è il suo territorio di caccia e che non desidera conspecifici estranei. La maggior parte dei canti emessi dagli uccelli servono a segnalare il possesso del territorio. I maschi di alcune falene possono emettere suoni con strutture microscopiche presenti nelle ali, e lo fanno durante la lotta per conquistarsi un luogo adatto per attrarre le femmine. Lo stesso vale per la stridulazione dei grilli e delle cicale che è usata per avvertire altri maschi del possesso del territorio. I grilli maschi sono molto aggressivi tra loro, infatti per allevarli e sentirli cantare è bene mantenerli in gabbie separate.
|
Gli insetti hanno sviluppato una grande varietà di adattamenti morfologici e comportamentali per evitare la predazione, tra cui anche la produzione di suoni. La stridulazione è usata oltre che per la comunicazione intraspecifica, talvolta anche a scopo difensivo antipredatorio. La specie che emette un segnale del genere, ha probabilmente armi temibili o è ben protetto e l’effetto del suono è quello di scoraggiare l’attacco di un eventuale predatore. |
Segnali d’allarme |
 Un segnale d’allarme viene emesso quando un animale intende comunicare ad un conspecifico la presenza di un pericolo imminente, come quello rappresentato dalla presenza di un predatore. Sono molto comuni tra i primati e gli uccelli. Il segnale per avere un valore comunicativo e adattativo, deve poter essere ricevuto e interpretato correttamente. Il ricevente deve poi decidere se e come rispondere. In genere un segnale d’allarme induce nel ricevente prima l’immobilizzazione, ma se il pericolo diventa più vicino segue la fuga. Il richiamo può anche variare le sue caratteristiche fisiche in funzione del tipo di predatore e della gravità del pericolo. Infatti, alcune scimmie sono in grado di emettere vocalizzazioni diverse in presenza di un’aquila o di un leopardo. Tali segnali specifici sono anche comprensibili per altre specie che ne possono usufruire. Infatti, si è notato che alcune specie di uccelli non rispondono agli allarmi delle scimmie che per loro non costituiscono un pericolo. Allo stesso modo, le cincie americane emettono segnali diversi in funzione delle dimensioni del predatore che anche in questo caso sono fruibili per altre specie. Molto studiati sono i segnali d’allarme degli scoiattoli. Interessante a questo proposito notare che essi sono l’unico caso sinora noto di specie che utilizzano gli ultrasuoni per comunicare il pericolo, pur utilizzando normalmente allo stesso scopo suoni nella gamma dell’udibile umano (fig. 17). Secondo alcuni autori l’ultrasonoro è usato in presenza di predatori incapaci di recepire in quelle frequenze. Un segnale d’allarme viene emesso quando un animale intende comunicare ad un conspecifico la presenza di un pericolo imminente, come quello rappresentato dalla presenza di un predatore. Sono molto comuni tra i primati e gli uccelli. Il segnale per avere un valore comunicativo e adattativo, deve poter essere ricevuto e interpretato correttamente. Il ricevente deve poi decidere se e come rispondere. In genere un segnale d’allarme induce nel ricevente prima l’immobilizzazione, ma se il pericolo diventa più vicino segue la fuga. Il richiamo può anche variare le sue caratteristiche fisiche in funzione del tipo di predatore e della gravità del pericolo. Infatti, alcune scimmie sono in grado di emettere vocalizzazioni diverse in presenza di un’aquila o di un leopardo. Tali segnali specifici sono anche comprensibili per altre specie che ne possono usufruire. Infatti, si è notato che alcune specie di uccelli non rispondono agli allarmi delle scimmie che per loro non costituiscono un pericolo. Allo stesso modo, le cincie americane emettono segnali diversi in funzione delle dimensioni del predatore che anche in questo caso sono fruibili per altre specie. Molto studiati sono i segnali d’allarme degli scoiattoli. Interessante a questo proposito notare che essi sono l’unico caso sinora noto di specie che utilizzano gli ultrasuoni per comunicare il pericolo, pur utilizzando normalmente allo stesso scopo suoni nella gamma dell’udibile umano (fig. 17). Secondo alcuni autori l’ultrasonoro è usato in presenza di predatori incapaci di recepire in quelle frequenze.
|
Richiami |
Il richiamo è messo in atto principalmente per attirare un conspecifico e principalmente dai maschi per attirare un potenziale partner. Sono, tuttavia, molto comuni anche tra i piccoli e le loro madri per ritrovarsi reciprocamente. Talvolta, il messaggio è indirizzato ad una specie diversa per facilitare un’eventuale collaborazione.
I suoni emessi sono caratteristici di ogni specie e talvolta sono discriminanti per il riconoscimento della specie, al fine di evitare improduttive ibridazione. E’ il caso di alcune specie di passeriformi e di rane, potenzialmente interfecondi, le cui femmine sono attratte solo dal canto del maschio della propria specie. Il richiamo sessuale è particolarmente importante quando gli individui di una specie sono poco sociali durante l’anno, ma devono necessariamente ritrovarsi nel periodo degli accoppiamenti. Ad esempio, molti felidi conducono vita solitaria e sono scarsamente o per niente loquaci, ma nel periodo degli amori emettono continui richiami. Ben noto è a tale proposito il fastidioso lamento notturno del gatto domestico. Nell’ambito dei richiami è interessante notare che nel babbuino femmina il verso ha un valore opposto. Esso viene emesso dopo l’accoppiamento con un maschio dominante e ha la funzione di tenere lontano altri maschi. |
 Gli insetti producono suoni mediante la vibrazione delle ali in volo e il ronzio prodotto per alcune specie ha valore comunicativo. Le diverse specie di zanzara femmine hanno una frequenza di vibrazione alare caratteristica, che serve al maschio per localizzare la femmina della sua specie (Fig. 18). Il frinire delle cicale e dei grilli oltre ad un ruolo di territorialità verso gli altri maschi, ha anche la funzione di richiamo sessuale per le femmine. Gli insetti producono suoni mediante la vibrazione delle ali in volo e il ronzio prodotto per alcune specie ha valore comunicativo. Le diverse specie di zanzara femmine hanno una frequenza di vibrazione alare caratteristica, che serve al maschio per localizzare la femmina della sua specie (Fig. 18). Il frinire delle cicale e dei grilli oltre ad un ruolo di territorialità verso gli altri maschi, ha anche la funzione di richiamo sessuale per le femmine.
 Molto studiati sotto l’aspetto etologico sono i versi emessi dai cuccioli degli animali domestici in difficoltà, i quali provocano una reazione immediata nella madre, che li afferra con la bocca e li porta al sicuro. Allo stesso modo la madre usa vocalizzazioni per richiamare a se i piccoli che si sono allontanati. Molto studiati sotto l’aspetto etologico sono i versi emessi dai cuccioli degli animali domestici in difficoltà, i quali provocano una reazione immediata nella madre, che li afferra con la bocca e li porta al sicuro. Allo stesso modo la madre usa vocalizzazioni per richiamare a se i piccoli che si sono allontanati.
|
L’emissione di suoni talvolta è utilizzata anche per richiami interspecifici. Infatti, alcune specie di bruchi sono in grado di emettere un ronzio di frequenza molto bassa, ma percepibile anche dall’orecchio umano, che attira le formiche della specie ecofilla smeraldina. Esse normalmente uccidono i bruchi a scopo alimentare, ma in questo caso il suono emesso comunica alla formica che il bruco è più utile da vivo, perché produce sostanze zuccherine di cui la formica è molto ghiotta, mediante specifiche ghiandole dorsali. Quindi, viene portato nel formicaio e allevato con cura.
L’uomo ha imparato a comunicare con le diverse specie domestiche, ma talvolta anche con specie selvatiche. Tra queste quella più spettacolare è l’indicatore dalla gola nera in Kenya, che viene richiamato con dei fischi caratteristici. L’indicatore emetterà anch’esso dei richiami per entrare in contatto con l’uomo; dopodiché lo guiderà verso un alveare da cui l’uomo ricaverà il miele. Per ricambiare il servizio offerto il favo viene aperto per lasciare l’accesso alle larve per l’indicatore. |
Richiesta di cibo |
Uno delle maggiori motivazioni che giustificano l’emissione di segnali sonori nelle specie che hanno cure parentali è la richiesta di cibo. I giovani di solito tendono ad acquisire rapidamente i segnali prodotti dagli adulti, tuttavia, continuano ad utilizzare per lungo tempo la gamma dei suoni giovanili diretti al genitore per richiedere cibo. I pulcini dei nidiacei sin dalla schiusa sono in grado di emettere pigolii non appena avvertono la presenza del genitore, in modo da stimolare l’imbeccata. In molti mammiferi e uccelli la vocalizzazione giovanile induce il rigurgito del cibo, che può essere utilizzato dai piccoli.
La richiesta di cibo è la vocalizzazione più tipica degli animali domestici. Peraltro, essi sono, per così dire, più loquaci dei loro consimili selvatici. Ciò è reso possibile dal fatto che vivono in un ambiente protetto, dove hanno imparato che farsi sentire non è affatto negativo: in questo contesto ambientale non ci sono ne predatori a cui sfuggire, ne prede da catturare. |
Ecolocalizzazione |
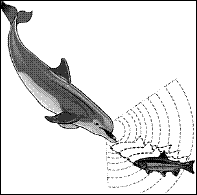 Uno dei fenomeni più spettacolari di utilizzo dei suoni in natura è l’ecolocalizzazione. Si tratta di un evento attuato da alcuni animali per rilevare informazioni spaziali analizzando i suoni che essi stessi emettono e che ritornano al loro orecchio, dopo essere stati riflessi dagli ostacoli. Gli animali che utilizzano questo sistema sono in grado di ottenere una mappa spaziale piuttosto precisa dell’ambiente in cui si muovono e di localizzare altri organismi viventi, come ad esempio le loro prede. Il gioco generalmente consiste nel calcolare il tempo che un suono impiega a ritornare alla sorgente per conoscerne la distanza. Però i sistemi di ecolocalizzazione più evoluti sono in grado di fare di più. L’emissione dei suoni ad altissima frequenza colpisce un ostacolo su tutta la sua superficie che quasi sempre non è piana, cioè presenta parti più vicine e altre più lontane dalla sorgente sonora, così alcune onde ritorneranno prima altre dopo. Il cervello, sulla base del diverso tempo di ritorno dell’onda, è in grado di ricostruire la struttura tridimensionale dell’ostacolo, estrapolando una mappa simile a quella che si ottiene attraverso la vista. Un sistema del genere è ovviamente alternativo a quello visivo in ambienti in cui la luce è limitata o di notte. Così è attuato dai pipistrelli che hanno abitudini notturne e si rifugiano in grotte buie e da alcuni uccelli, come le salangane, che nidificano in caverne. Inoltre, è anche utilizzato dai cetacei (Fig. 19), che si muovono in un ambiente in cui la luce si estingue rapidamente dopo qualche centinaio di metri di profondità. In queste circostanze i suoni sono molto più utili rispetto agli altri sensi, e non solo ai fini dell’ecolocalizzazione, ma anche perchè si muovono velocemente, su grandi distanze e possono essere direzionati dove più servono. Secondo alcuni ricercatori il canto delle megattere servirebbe a localizzare a distanza gli individui della stessa specie. Alcuni delfini di fiume sono del tutto ciechi e, quindi, si orientano esclusivamente attraverso il biosonar. Ben noto è a tal proposito il delfino del Gange, un ambiente ricco di particelle inorganiche in sospensione che riducono il campo visivo a qualche metro. Alcuni studi sui delfini che vivono in mare hanno evidenziato che essi apprendono durante la loro vita, probabilmente già nella fase embrionale dalla madre, ad utilizzare correttamente il biosonar. Cioè c’è una componente appresa che, peraltro è caratteristica del guppo. In pratica, una comunità di delfini che interagiscono tra loro utilizza il biosonar in maniera un po’ diversa rispetto a delfini con cui non si hanno relazioni. E’ come se ci fossero dei dialetti locali nell’uso dei segnali acustici ad alta frequenza. Le vocalizzazioni che sono invece utilizzate per le normali conversazioni, sono invece innate e sono invariabili per tutti i delfini di una stessa specie. Uno dei fenomeni più spettacolari di utilizzo dei suoni in natura è l’ecolocalizzazione. Si tratta di un evento attuato da alcuni animali per rilevare informazioni spaziali analizzando i suoni che essi stessi emettono e che ritornano al loro orecchio, dopo essere stati riflessi dagli ostacoli. Gli animali che utilizzano questo sistema sono in grado di ottenere una mappa spaziale piuttosto precisa dell’ambiente in cui si muovono e di localizzare altri organismi viventi, come ad esempio le loro prede. Il gioco generalmente consiste nel calcolare il tempo che un suono impiega a ritornare alla sorgente per conoscerne la distanza. Però i sistemi di ecolocalizzazione più evoluti sono in grado di fare di più. L’emissione dei suoni ad altissima frequenza colpisce un ostacolo su tutta la sua superficie che quasi sempre non è piana, cioè presenta parti più vicine e altre più lontane dalla sorgente sonora, così alcune onde ritorneranno prima altre dopo. Il cervello, sulla base del diverso tempo di ritorno dell’onda, è in grado di ricostruire la struttura tridimensionale dell’ostacolo, estrapolando una mappa simile a quella che si ottiene attraverso la vista. Un sistema del genere è ovviamente alternativo a quello visivo in ambienti in cui la luce è limitata o di notte. Così è attuato dai pipistrelli che hanno abitudini notturne e si rifugiano in grotte buie e da alcuni uccelli, come le salangane, che nidificano in caverne. Inoltre, è anche utilizzato dai cetacei (Fig. 19), che si muovono in un ambiente in cui la luce si estingue rapidamente dopo qualche centinaio di metri di profondità. In queste circostanze i suoni sono molto più utili rispetto agli altri sensi, e non solo ai fini dell’ecolocalizzazione, ma anche perchè si muovono velocemente, su grandi distanze e possono essere direzionati dove più servono. Secondo alcuni ricercatori il canto delle megattere servirebbe a localizzare a distanza gli individui della stessa specie. Alcuni delfini di fiume sono del tutto ciechi e, quindi, si orientano esclusivamente attraverso il biosonar. Ben noto è a tal proposito il delfino del Gange, un ambiente ricco di particelle inorganiche in sospensione che riducono il campo visivo a qualche metro. Alcuni studi sui delfini che vivono in mare hanno evidenziato che essi apprendono durante la loro vita, probabilmente già nella fase embrionale dalla madre, ad utilizzare correttamente il biosonar. Cioè c’è una componente appresa che, peraltro è caratteristica del guppo. In pratica, una comunità di delfini che interagiscono tra loro utilizza il biosonar in maniera un po’ diversa rispetto a delfini con cui non si hanno relazioni. E’ come se ci fossero dei dialetti locali nell’uso dei segnali acustici ad alta frequenza. Le vocalizzazioni che sono invece utilizzate per le normali conversazioni, sono invece innate e sono invariabili per tutti i delfini di una stessa specie.
|
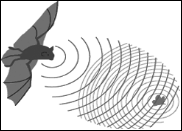 I pipistrelli sono in grado di memorizzare la mappa spaziale ottenuta attraverso il biosonar, ciò che consente loro di compiere i percorsi abituali senza utilizzare il sistema di ecolocalizzazione, evitando, così, un inutile spreco di energia; un po’ come fanno le persone cieche che hanno memorizzato gli ostacoli della loro casa. Per contro l’emissione dei segnali acustici emessi aumenta notevolmente una volta localizzata una possibile preda. I pipistrelli possono anche ricavare informazioni sulla direzione di movimento della preda dalla differenza infinitesimale di tempo con cui le onde sonore colpiscono le due orecchie (fig. 20). I pipistrelli sono in grado di memorizzare la mappa spaziale ottenuta attraverso il biosonar, ciò che consente loro di compiere i percorsi abituali senza utilizzare il sistema di ecolocalizzazione, evitando, così, un inutile spreco di energia; un po’ come fanno le persone cieche che hanno memorizzato gli ostacoli della loro casa. Per contro l’emissione dei segnali acustici emessi aumenta notevolmente una volta localizzata una possibile preda. I pipistrelli possono anche ricavare informazioni sulla direzione di movimento della preda dalla differenza infinitesimale di tempo con cui le onde sonore colpiscono le due orecchie (fig. 20).
|
 Inoltre, sono anche capaci di capire se si stanno avvicinando alla preda o allontanando, mediante l’analisi dell’effetto Doppler sulle onde sonore. Il suono emesso da una sorgente in movimento sono avvertite con una frequenza maggiore quando si avvicinano e minore quando si allontanano (fig. 21). Inoltre, sono anche capaci di capire se si stanno avvicinando alla preda o allontanando, mediante l’analisi dell’effetto Doppler sulle onde sonore. Il suono emesso da una sorgente in movimento sono avvertite con una frequenza maggiore quando si avvicinano e minore quando si allontanano (fig. 21).
|
 Questo è il motivo per cui un treno che passa davanti ad un ascoltatore varia la sua frequenza del rumore. E’ come se ci fosse una compressione delle onde se la sorgente si muove nella direzione dell’ascoltatore, per cui un maggior numero di onde per unità di tempo e, quindi, una maggiore frequenza. L’analisi delle frequenze degli ultrasuoni di ritorno consente quindi al pipistrello di capire se si sta avvicinando alla preda e di prevederne il contatto (fig. 22). Si è anche osservato che il segnale emesso non ha una frequenza fissa, ma può essere modulato. Non si sa bene perché, ma s’ipotizza che questo serva per ricevere dati anche qualitativi, per esempio capire il tipo di preda. Questo è il motivo per cui un treno che passa davanti ad un ascoltatore varia la sua frequenza del rumore. E’ come se ci fosse una compressione delle onde se la sorgente si muove nella direzione dell’ascoltatore, per cui un maggior numero di onde per unità di tempo e, quindi, una maggiore frequenza. L’analisi delle frequenze degli ultrasuoni di ritorno consente quindi al pipistrello di capire se si sta avvicinando alla preda e di prevederne il contatto (fig. 22). Si è anche osservato che il segnale emesso non ha una frequenza fissa, ma può essere modulato. Non si sa bene perché, ma s’ipotizza che questo serva per ricevere dati anche qualitativi, per esempio capire il tipo di preda.
I pipistrelli possono arrivare ad emettere anche 200 impulsi al secondo a frequenza elevatissima. Molto meno capaci sono gli uccelli del genere Aerodramus, per cui il loro biosonar risulta meno efficace. Questi uccelli nidificano nella profondità delle grotte dove la luce non arriva, così emettono una serie d’impulsi sonori udibili per l’uomo (tra 2000 e 8000 Hz) che s’intensificano quando arrivano in prossimità del posatoio, fino al massimo di circa 30 al secondo e a frequenze meno elevate rispetto ai pipistrelli. Ciò si traduce in una mappa spaziale molto meno dettagliata.
|
Organi per la produzione dei suoni |
 I suoni in natura possono essere emessi in maniera molto diversa. Alcuni animali sono in grado di produrre suoni con valore comunicativo anche in assenza di organi specifici. Vi sono specie che colpiscono su substrati di diverso genere con parti del corpo. I picchi battono con il becco contro le cortecce provocando un tambureggiamento tipico di ogni singola specie; questo comportamento ha la funzione di segnalare il possesso del territorio (Fig. 23). Gli spalaci, che somigliano alle nostre talpe per quanto attiene alle loro abitudini, scavano gallerie sotto il terreno; essi battono spesso con la testa sotto il soffitto del tunnel per indicare la propria posizione ad un conspecifico ed evitare, così, un pericoloso scontro fisico, che potrebbe essere fatale. I suoni in natura possono essere emessi in maniera molto diversa. Alcuni animali sono in grado di produrre suoni con valore comunicativo anche in assenza di organi specifici. Vi sono specie che colpiscono su substrati di diverso genere con parti del corpo. I picchi battono con il becco contro le cortecce provocando un tambureggiamento tipico di ogni singola specie; questo comportamento ha la funzione di segnalare il possesso del territorio (Fig. 23). Gli spalaci, che somigliano alle nostre talpe per quanto attiene alle loro abitudini, scavano gallerie sotto il terreno; essi battono spesso con la testa sotto il soffitto del tunnel per indicare la propria posizione ad un conspecifico ed evitare, così, un pericoloso scontro fisico, che potrebbe essere fatale.
|
Anche i colombi che posseggono un organo ben strutturato per la produzione dei suoni usano spesso sorgenti alternative. Essi battono con forza le ali facendole urtare superiormente tra loro; in questo modo emettono una specie di schiocco che segnala la presenza di un pericolo. Analogamente, alcuni fagiani delle foreste tropicali battono con vigore le ali quando vogliono comunicare il possesso del territorio. Noi stessi con l’applauso intendiamo comunicare il gradimento e gratificare l’artista. Alcuni pesci utilizzano i denti, che strofinando tra loro producono suono stridulo; altri pesci possono emettere suoni espellendo rapidamente gas dalla bocca. Tuttavia, in questi vertebrati il modo più comune per produrre dei suoni è mediante la vibrazione della vescica natatoria (Fig. 24). In alcuni casi quest’organo possiede dei muscoli specifici che, nel rospo marino, lo fanno vibrare fino a 200 volte al secondo. Un altro modo del tutto caratteristico di emettere suoni è quello del crotalo, che presenta all’estremità della coda una specie di sonaglio, che viene fatto vibrare per segnalare la posizione  taglia. L’organo è costituito da lembi di pelle che dopo la muta, anziché essere eliminati, si arrotolano posteriormente formando il sonaglio. taglia. L’organo è costituito da lembi di pelle che dopo la muta, anziché essere eliminati, si arrotolano posteriormente formando il sonaglio. |
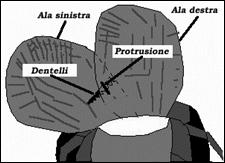 Molti insetti possiedono organi specifici per la produzione di suoni, che si sono evoluti a partire da elementi strutturali diversi. Gli ortotteri posseggono delle ali anteriori coriacee e poco adatte al volo, dette tegmine, le quali sono coinvolte nella produzione del suono. Esse in alcune specie di cavalletta sono sfregate con le ali posteriori; in certe altre specie, invece, con il femore delle zampe posteriori. In questo caso i femori presentano dei dentelli o asperità che facilitano la stridulazione. Nei grilli le tegmine sono semplicemente sfregate tra di loro (fig. 25); in questo caso la termina destra e quella sinistra sono spesso strutturate in maniera diversa, in maniera tale da ottimizzare lo stridio. I coleotteri sfregano tra loro pezzi dell’esoscheletro toracico e alcuni emitteri parti boccali sull’esoscheletro basale. Nelle cicale gli organi per la produzione dei suoni si trovano lateralmente al torace. Essi sono costituiti da cavità sul fondo delle quali è presente una membrana connessa a dei muscoli. La contrazione coordinata dei muscoli fa variare rapidamente la concavità della membrana, il che produce delle vibrazioni sonore, le quali si amplificano nella cavità. Molti insetti possiedono organi specifici per la produzione di suoni, che si sono evoluti a partire da elementi strutturali diversi. Gli ortotteri posseggono delle ali anteriori coriacee e poco adatte al volo, dette tegmine, le quali sono coinvolte nella produzione del suono. Esse in alcune specie di cavalletta sono sfregate con le ali posteriori; in certe altre specie, invece, con il femore delle zampe posteriori. In questo caso i femori presentano dei dentelli o asperità che facilitano la stridulazione. Nei grilli le tegmine sono semplicemente sfregate tra di loro (fig. 25); in questo caso la termina destra e quella sinistra sono spesso strutturate in maniera diversa, in maniera tale da ottimizzare lo stridio. I coleotteri sfregano tra loro pezzi dell’esoscheletro toracico e alcuni emitteri parti boccali sull’esoscheletro basale. Nelle cicale gli organi per la produzione dei suoni si trovano lateralmente al torace. Essi sono costituiti da cavità sul fondo delle quali è presente una membrana connessa a dei muscoli. La contrazione coordinata dei muscoli fa variare rapidamente la concavità della membrana, il che produce delle vibrazioni sonore, le quali si amplificano nella cavità.
|
La maggioranza dei vertebrati utilizza per la produzione dei suoni le vie aeree, dove si localizzano organi specifici. Il suono può uscire dalla bocca oppure dal naso. Talvolta sono presenti degli organi aggiuntivi, che oltre alla produzione servono anche per direzionare e amplificare i suoni. Nelle rane ad esempio ci sono spesso dei sacchi vocali che possono essere localizzati ai lati della bocca (rane verdi) o sotto la gola (raganelle) che consentono una forte amplificazione del segnale sonoro. D’altra parte nei mammiferi le cavità della bocca e quelle nasali hanno generalmente un effetto amplificante. |
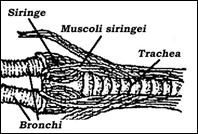 Negli uccelli, tranne che in alcuni rari casi (es. cicogne, struzzi), c’è la siringe. Si tratta di un organo situato alla congiunzione tra la trachea e i bronchi, che l’animale può far vibrare in maniera diversa modulando la forza con cui emette l’aria. La siringe in alcune specie appare piuttosto semplice, mentre in altre è complicata da una serie di membrane ed elementi muscolari (Fig. 26). Negli uccelli, tranne che in alcuni rari casi (es. cicogne, struzzi), c’è la siringe. Si tratta di un organo situato alla congiunzione tra la trachea e i bronchi, che l’animale può far vibrare in maniera diversa modulando la forza con cui emette l’aria. La siringe in alcune specie appare piuttosto semplice, mentre in altre è complicata da una serie di membrane ed elementi muscolari (Fig. 26).
|
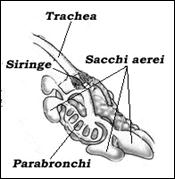 Al meccanismo di fonazione degli uccelli partecipano in maniera determinante i sacchi aerei, i quali, grazie all’effetto risonanza amplificano l’intensità dell’emissione sonora (Fig. 27). Al meccanismo di fonazione degli uccelli partecipano in maniera determinante i sacchi aerei, i quali, grazie all’effetto risonanza amplificano l’intensità dell’emissione sonora (Fig. 27).
 Come negli uccelli anche nei mammiferi c’è un elaborato organo costituito da una serie di membrane dette corde vocali. Le corde vocali, diversamente dalla siringe sono allogate più in alto nelle vie respiratorie, in prossimità della laringe (Fig. 28). Esse sono strutture lamellari costituite da uno scheletro cartilagineo, muscoli e legamenti. I muscoli sono responsabili dello stato di contrazione dell’organo, che a sua volta determina il tipo vibrazione e, quindi, la frequenza del suono emesso. Nell’uomo le corde vocali del maschio risultano più lunghe rispetto a quelle della femmina, in media del 30%. Questo è il motivo per cui le donne sono in grado di raggiungere tonalità più elevate. L’intensità del suono invece dipende dalla velocità con cui il flusso d’aria passa sulle corde vocali. Al meccanismo di fonazione partecipano i muscoli del cavo orale e la lingua. Come negli uccelli anche nei mammiferi c’è un elaborato organo costituito da una serie di membrane dette corde vocali. Le corde vocali, diversamente dalla siringe sono allogate più in alto nelle vie respiratorie, in prossimità della laringe (Fig. 28). Esse sono strutture lamellari costituite da uno scheletro cartilagineo, muscoli e legamenti. I muscoli sono responsabili dello stato di contrazione dell’organo, che a sua volta determina il tipo vibrazione e, quindi, la frequenza del suono emesso. Nell’uomo le corde vocali del maschio risultano più lunghe rispetto a quelle della femmina, in media del 30%. Questo è il motivo per cui le donne sono in grado di raggiungere tonalità più elevate. L’intensità del suono invece dipende dalla velocità con cui il flusso d’aria passa sulle corde vocali. Al meccanismo di fonazione partecipano i muscoli del cavo orale e la lingua.
|
 Nei vertebrati il suono è generalmente emesso dalla bocca, tuttavia può anche essere emesso dal naso. A tale proposito i pipistrelli della famiglia dei rinolofi, presentano una struttura carnosa rilevata in prossimità del naso, detta foglia nasale, caratteristica di ogni specie, che serve a direzionare i suoni nasali. Alcune strutture aggiuntive che ottimizzano la produzione del suono sono anche presenti nei cetacei. Negli odontoceti è presente il melone, una formazione adiposa localizzata anteriormente all’osso frontale, che ha la funzione di trasmettere e direzionare i suoni prodotti in un complesso sistema di sacchi aerei presenti nelle vie nasali. Il melone può essere più o meno esteso e, pertanto, modella il capo delle diverse specie di delfini in maniera del tutto caratteristica (Fig. 29). In altri casi, come nel capodoglio, al posto del melone è presente lo spermaceti, costituito da muscoli che circondano un tessuto adiposo, che, a sua volta, racchiude un liquido denso. Questo liquido può cambiare le sue caratteristiche fisiche con il variare dal profondità e la temperatura; tuttavia, il suo coinvolgimento nel meccanismo di modifica e direzionamento dei suoni non e ancora del tutto certo. Nei vertebrati il suono è generalmente emesso dalla bocca, tuttavia può anche essere emesso dal naso. A tale proposito i pipistrelli della famiglia dei rinolofi, presentano una struttura carnosa rilevata in prossimità del naso, detta foglia nasale, caratteristica di ogni specie, che serve a direzionare i suoni nasali. Alcune strutture aggiuntive che ottimizzano la produzione del suono sono anche presenti nei cetacei. Negli odontoceti è presente il melone, una formazione adiposa localizzata anteriormente all’osso frontale, che ha la funzione di trasmettere e direzionare i suoni prodotti in un complesso sistema di sacchi aerei presenti nelle vie nasali. Il melone può essere più o meno esteso e, pertanto, modella il capo delle diverse specie di delfini in maniera del tutto caratteristica (Fig. 29). In altri casi, come nel capodoglio, al posto del melone è presente lo spermaceti, costituito da muscoli che circondano un tessuto adiposo, che, a sua volta, racchiude un liquido denso. Questo liquido può cambiare le sue caratteristiche fisiche con il variare dal profondità e la temperatura; tuttavia, il suo coinvolgimento nel meccanismo di modifica e direzionamento dei suoni non e ancora del tutto certo.
|
Organi per la percezione dei suoni |
la maggior parte dei phyla animali, fatta eccezione per gli artropodi e i cordati, è costituito da specie incapaci di produrre e percepire i suoni. La presenza di organi adatti per la ricezione di segnali sonori è un’ovvia presenza nelle specie che comunicano attraverso i suoni, ma essi sono anche presenti in specie mute. |
Insetti |
 Tra gli artropodi i più noti comunicatori acustistici appartengono al gruppo degli insetti. La percezione dei suoni avviene attraverso delle cellule sensoriali disperse sul corpo, gli scolopidii, che spesso si associano tra di loro in gruppetti per formare organi più complessi, detti scolopofori. Questi elementi cellulari sono parte integrante degli organi cordonali, che sono strutture allungate che percepiscono il suono vibrando. Essi possono essere presenti in varie parti del corpo, come ad esempio le antenne (fig. 30). Tra gli artropodi i più noti comunicatori acustistici appartengono al gruppo degli insetti. La percezione dei suoni avviene attraverso delle cellule sensoriali disperse sul corpo, gli scolopidii, che spesso si associano tra di loro in gruppetti per formare organi più complessi, detti scolopofori. Questi elementi cellulari sono parte integrante degli organi cordonali, che sono strutture allungate che percepiscono il suono vibrando. Essi possono essere presenti in varie parti del corpo, come ad esempio le antenne (fig. 30).
|
A parte gli organi cordonali in alcuni insetti è presente una vera e propria membrana timpanica. Nei grilli il timpano si trova sulle zampe anteriori, nelle cicale è addominale e nelle falene della famiglia dei Noctuidi il è localizzato lateralmente al torace. Queste farfalle notturne non comunicano tra loro attraverso suoni e la loro membrana timpanica è in grado di vibrare sotto lo stimolo degli ultrasuoni dei pipistrelli. In questo modo sono in grado di mettere in atto comportamenti diversivi per evitare di essere predate. |
Vertebrati |
Tra i cordati solo i vertebrati sono hanno organi per percepire i suoni. La componente comune a tutti i gruppi è l’orecchio, ma nei pesci e negli anfibi larvali è presente una struttura aggiuntiva, la linea laterale. |
L’orecchio |
 L’orecchio è articolato in maniera diversa nelle varie specie, ma sostanzialmente risponde ad un unico palinsesto strutturale. Nei pesci è situato dietro la testa ed è incapsulato in una struttura cartilaginea od ossea. Esso è costituito da tre canali perpendicolari tra loro (canali semicircolari) che confluiscono in un canale più largo (sacculo). Questi elementi non partecipano alla percezione del suono, ma sono organi dell’equilibrio. Dal sacculo si diparte un breve canale, la lagena, che contiene l’organo per la decodifica dei suoni. Tutti questi elementi costituiscono nel complesso l’orecchio interno. Nei pesci non è presente una membrana timpanica, tuttavia, spesso la vescica natatoria può fungere da cassa di risonanza per amplificare i suoni in arrivo. In alcuni casi sono anche presenti degli ossicoli (ossicini di Weber) che trasmettono il suono dalla vescica natatoria alla lagena (Fig. 31). Gli ossicini di Weber non hanno alcuna relazione evolutiva con quelli dell’orecchio dei tetrapodi e, pertanto, non devono essere considerati come parte dell’orecchio medio, che, invece, è una porzione del sistema uditivo che ritroviamo a partire dagli anfibi. L’orecchio è articolato in maniera diversa nelle varie specie, ma sostanzialmente risponde ad un unico palinsesto strutturale. Nei pesci è situato dietro la testa ed è incapsulato in una struttura cartilaginea od ossea. Esso è costituito da tre canali perpendicolari tra loro (canali semicircolari) che confluiscono in un canale più largo (sacculo). Questi elementi non partecipano alla percezione del suono, ma sono organi dell’equilibrio. Dal sacculo si diparte un breve canale, la lagena, che contiene l’organo per la decodifica dei suoni. Tutti questi elementi costituiscono nel complesso l’orecchio interno. Nei pesci non è presente una membrana timpanica, tuttavia, spesso la vescica natatoria può fungere da cassa di risonanza per amplificare i suoni in arrivo. In alcuni casi sono anche presenti degli ossicoli (ossicini di Weber) che trasmettono il suono dalla vescica natatoria alla lagena (Fig. 31). Gli ossicini di Weber non hanno alcuna relazione evolutiva con quelli dell’orecchio dei tetrapodi e, pertanto, non devono essere considerati come parte dell’orecchio medio, che, invece, è una porzione del sistema uditivo che ritroviamo a partire dagli anfibi.
|
L’orecchio medio presenta un osso, la staffa, che amplifica il segnale percepito. Nei mammiferi, oltre alla staffa, vi sono altri due ossicini, l’incudine e il martello. Essi derivano da ossa dell’articolazione mandibolare dei rettili (in particolare, dal quadrato e l’articolare) e s’interpongono tra la staffa e il timpano. L’amplificazione del suono nell’orecchio medio è molto efficiente; cosicché suoni eccessivamente intensi possono danneggiare le strutture auricolari interne. A tale proposito, i pipistrelli che emettono suoni di grande intensità nel meccanismo di ecolocalizzazione, hanno muscoli molto efficaci nel disarticolare, molte volte al secondo, gli ossicini nell’istante in cui viene emesso l’impulso. Meccanismi del genere avvengono anche nei mammiferi, ma con minore efficacia, in quanto i muscoli reagiscono all’impulso con un certo ritardo e, quindi, non proteggono l’orecchio da rumori improvvisi, come quello dello scoppio di una bomba. Inoltre, funzionano solo a basse frequenze. La membrana timpanica segna il limite tra l’orecchio esterno e quello medio, mentre il segnale amplificato dagli ossicini viene trasferito ad un’altra membrana, la finestra ovale, che, invece, separa l’orecchio medio da quello interno. Diversamente dall’orecchio medio dei tetrapodi, quello interno corrisponde evolutivamente a quello dei pesci; infatti, ritroviamo i canali semicircolari e la lagena, anche se, in ogni specie, strutturati in maniera un po’ diversa. |
 Nella maggior parte dei casi esiste anche un orecchio esterno formato da condotto uditivo, che convoglia i suoni verso la membrana timpanica, situata sul fondo del canale. L’orecchio esterno è assente negli anfibi, e pertanto il loro timpano è visibile ai lati del capo. Nei mammiferi, invece, il condotto uditivo è circondato esternamente da un padiglione auricolare utile a canalizzare meglio i suoni (Fig. 32). Questa struttura può essere opportunamente direzionata mediante dei muscoli specifici controllati volontariamente, i muscoli auricolari. Essi sono presenti anche nell’uomo, tuttavia, solo pochi individui sono ancora in grado di farli contrarre muovendo lievemente le orecchie. Nella maggior parte dei casi esiste anche un orecchio esterno formato da condotto uditivo, che convoglia i suoni verso la membrana timpanica, situata sul fondo del canale. L’orecchio esterno è assente negli anfibi, e pertanto il loro timpano è visibile ai lati del capo. Nei mammiferi, invece, il condotto uditivo è circondato esternamente da un padiglione auricolare utile a canalizzare meglio i suoni (Fig. 32). Questa struttura può essere opportunamente direzionata mediante dei muscoli specifici controllati volontariamente, i muscoli auricolari. Essi sono presenti anche nell’uomo, tuttavia, solo pochi individui sono ancora in grado di farli contrarre muovendo lievemente le orecchie.
|
L’organo della linea laterale |
 Questa struttura è costituita da una serie di canali che decorrono lateralmente al corpo del pesce in maniera rettilinea, ma in prossimità della testa si complicano, articolandosi in una serie di diramazioni tutte in collegamento tra loro. Questi canali sono in collegamento con l’esterno mediante una serie di pori che consentono l’entrata e l’uscita dell’acqua (Fig. 33). All’interno dei canali, sporgono una serie di gruppetti cellulari dotati di ciglia sensoriali ricoperte e agglutinate da una sostanza gelatinosa. Questa struttura è il neuromasto che, come i peli sensoriali degli insetti, non rileva le onde di pressione, ma il movimento delle particelle. La linea laterale consente di percepire suoni con frequenza molto bassa (circa 100 Hz), mentre l’orecchio del pesce è adatto per suoni con maggiore frequenza (superiori a 1000 Hz). Oltre che a percepire i suoni l’organo consente ai pesci di passare in spazi angusti velocemente senza toccare il substrato ed evitando di ferirsi. Ciò è possibile in quanto lo stesso movimento del pesce provoca delle onde meccaniche che rimbalzano sulle rocce e ritornano alla linea laterale informando, il pesce sulla distanza dell’ostacolo situato lateralmente, e quindi al di fuori del campo visivo. La linea laterale è anche utile per avvertire l’arrivo di un predatore con il necessario anticipo. Un animale in movimento genera delle onde meccaniche che viaggiano più veloci di lui e, quindi, arrivano alla linea laterale di una sua potenziale preda prima; ciò che consente di mettere in atto una manovra diversiva. La linea laterale è presente anche nei girini delle rane, ma scompare dopo la metamorfosi. Questa struttura è costituita da una serie di canali che decorrono lateralmente al corpo del pesce in maniera rettilinea, ma in prossimità della testa si complicano, articolandosi in una serie di diramazioni tutte in collegamento tra loro. Questi canali sono in collegamento con l’esterno mediante una serie di pori che consentono l’entrata e l’uscita dell’acqua (Fig. 33). All’interno dei canali, sporgono una serie di gruppetti cellulari dotati di ciglia sensoriali ricoperte e agglutinate da una sostanza gelatinosa. Questa struttura è il neuromasto che, come i peli sensoriali degli insetti, non rileva le onde di pressione, ma il movimento delle particelle. La linea laterale consente di percepire suoni con frequenza molto bassa (circa 100 Hz), mentre l’orecchio del pesce è adatto per suoni con maggiore frequenza (superiori a 1000 Hz). Oltre che a percepire i suoni l’organo consente ai pesci di passare in spazi angusti velocemente senza toccare il substrato ed evitando di ferirsi. Ciò è possibile in quanto lo stesso movimento del pesce provoca delle onde meccaniche che rimbalzano sulle rocce e ritornano alla linea laterale informando, il pesce sulla distanza dell’ostacolo situato lateralmente, e quindi al di fuori del campo visivo. La linea laterale è anche utile per avvertire l’arrivo di un predatore con il necessario anticipo. Un animale in movimento genera delle onde meccaniche che viaggiano più veloci di lui e, quindi, arrivano alla linea laterale di una sua potenziale preda prima; ciò che consente di mettere in atto una manovra diversiva. La linea laterale è presente anche nei girini delle rane, ma scompare dopo la metamorfosi.
|
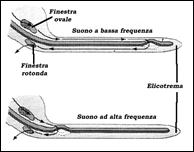
 della chiocciola, cioè nel punto più distante dalla finestra ovale, tutte le membrane s’interrompono, per cui i canali longitudinali entrano in comunicazione in un’area detta elicotrema. della chiocciola, cioè nel punto più distante dalla finestra ovale, tutte le membrane s’interrompono, per cui i canali longitudinali entrano in comunicazione in un’area detta elicotrema.
|
La percezione nell’orecchio esterno |
 Un segnale qualsiasi sonoro è convogliato al timpano mediante l’orecchio esterno (qualora presente) in maniera tale da essere più o meno amplificato. Cioè l’orecchio esterno non solo recepisce il messaggio sonoro favorendone l’intercettazione spaziale mediante un eventuale padiglione auricolare, ma riesce anche ad aumentarne l’ampiezza (Fig. 36). Nell’uomo grazie all’orecchio esterno la pressione sonora che arriva al timpano può essere anche 10 volte maggiore di quella del segnale originario. Per un fatto puramente strutturale del padiglione auricolare, nell’uomo l’effetto amplificante è più efficace nelle frequenze comprese tra 2500 e 5000 Hz. Un segnale qualsiasi sonoro è convogliato al timpano mediante l’orecchio esterno (qualora presente) in maniera tale da essere più o meno amplificato. Cioè l’orecchio esterno non solo recepisce il messaggio sonoro favorendone l’intercettazione spaziale mediante un eventuale padiglione auricolare, ma riesce anche ad aumentarne l’ampiezza (Fig. 36). Nell’uomo grazie all’orecchio esterno la pressione sonora che arriva al timpano può essere anche 10 volte maggiore di quella del segnale originario. Per un fatto puramente strutturale del padiglione auricolare, nell’uomo l’effetto amplificante è più efficace nelle frequenze comprese tra 2500 e 5000 Hz.
|
La trasmissione nell’orecchio medio |
 Il timpano vibra trasmettendo il segnale agli ossicini dell’orecchio medio; nell’ordine martello, incudine e staffa (Fig 37). A questo livello il suono è trasformato in un messaggio meccanico con un certo contenuto di energia. Questo messaggio è trasferito al liquido cocleare mediante l’azione della staffa sulla finestra ovale. La catena di ossicini non ha però solo la funzione di trasferimento dell’energia acustica, ma anche quella di amplificarla. Ciò si spiega se pensiamo che gli ossicini sono articolati mediante dei meccanismi di leva molto favorevoli; d’altra parte il timpano risulta molto più grande della finestra ovale e, quindi, l’energia recepita dal timpano alla fine si distribuisce sulla minore superficie della finestra ovale, concentrandosi. Nell’uomo è stato calcolato che la pressione esercitata dalla staffa sulla finestra ovale è di circa 185 volte maggiore rispetto a quella esercitata dal suono sul timpano. E’ stato anche osservato che gli ossicini sono meglio adattati a trasmettere i suoni con maggiore efficacia per frequenze comprese tra 1000 e 2000 Hz. Forme di otiti possono provocare l’immobilità del timpano, il che determina una minore capacità di trasmissione dei suoni. Allo stesso modo proliferazioni ossee nell’orecchio medio possono ridurre o impedire la funzionalità degli ossicini. Entrambi i casi possono portare alla sordità totale. Il timpano vibra trasmettendo il segnale agli ossicini dell’orecchio medio; nell’ordine martello, incudine e staffa (Fig 37). A questo livello il suono è trasformato in un messaggio meccanico con un certo contenuto di energia. Questo messaggio è trasferito al liquido cocleare mediante l’azione della staffa sulla finestra ovale. La catena di ossicini non ha però solo la funzione di trasferimento dell’energia acustica, ma anche quella di amplificarla. Ciò si spiega se pensiamo che gli ossicini sono articolati mediante dei meccanismi di leva molto favorevoli; d’altra parte il timpano risulta molto più grande della finestra ovale e, quindi, l’energia recepita dal timpano alla fine si distribuisce sulla minore superficie della finestra ovale, concentrandosi. Nell’uomo è stato calcolato che la pressione esercitata dalla staffa sulla finestra ovale è di circa 185 volte maggiore rispetto a quella esercitata dal suono sul timpano. E’ stato anche osservato che gli ossicini sono meglio adattati a trasmettere i suoni con maggiore efficacia per frequenze comprese tra 1000 e 2000 Hz. Forme di otiti possono provocare l’immobilità del timpano, il che determina una minore capacità di trasmissione dei suoni. Allo stesso modo proliferazioni ossee nell’orecchio medio possono ridurre o impedire la funzionalità degli ossicini. Entrambi i casi possono portare alla sordità totale.
|
La trasduzione nell’organo del Corti
 Il movimento del liquido cocleare in seguito all’azione della staffa mette in vibrazione la membrana basilare. La diversa disposizione delle fibre elastiche nella membrana basilare determina delle caratteristiche per cui la parte più vicina alla finestra ovale è più rigida, per divenire sempre più elastica procedendo verso l’apice della chiocciola. A causa di questa organizzazione strutturale le frequenze più alte determinano dei picchi di oscillazione e sono percepite all’inizio della chiocciola e quelle progressivamente più basse sempre più lontane (Fig. 38). In pratica, anche se la membrana basilare è più elastica verso l’apice della chiocciola, e un suono acuto potrebbe potenzialmente metterla in vibrazione, ciò in realtà non accade perché la sua energia si scarica prima. Il movimento del liquido cocleare in seguito all’azione della staffa mette in vibrazione la membrana basilare. La diversa disposizione delle fibre elastiche nella membrana basilare determina delle caratteristiche per cui la parte più vicina alla finestra ovale è più rigida, per divenire sempre più elastica procedendo verso l’apice della chiocciola. A causa di questa organizzazione strutturale le frequenze più alte determinano dei picchi di oscillazione e sono percepite all’inizio della chiocciola e quelle progressivamente più basse sempre più lontane (Fig. 38). In pratica, anche se la membrana basilare è più elastica verso l’apice della chiocciola, e un suono acuto potrebbe potenzialmente metterla in vibrazione, ciò in realtà non accade perché la sua energia si scarica prima. |
La finestra rotonda deformandosi è utile per assorbire la vibrazione; in caso contrario un segnale in arrivo potrebbe rimbalzare sulle superfici ossee provocando disturbi nella percezione dei segnali successivi.
L’oscillazione locale della membrana basilare in seguito ad una determinata frequenza può essere più o meno accentuata in funzione dell’intensità dello stimolo. Di conseguenza, suoni più forti provocano una maggiore vibrazione della membrana (nell’ambito di quella frequenza) e, quindi, sono percepiti con volumi più alti.
La vibrazione della membrana basilare mette in funzione la membrana tettoria, che inizialmente sposta le ciglia, a cui è legata strutturalmente, delle cellule sensoriali esterne. Ciò determina l’apertura di canali per particelle con cariche positive (cationi) che entrano nella cellula dall’ambiente circostante. Il loro movimento verso l’interno provoca una corrente che induce la contrazione delle cellule sensoriali esterne, accompagnate dalle cellule del Deiters sottostanti. La contrazione è poi è ritrasmessa alla membrana tettoria. Le fibre nervose connesse alle cellule esterne sono in larga parte afferenti, cioè sono fibre che conducono dei segnali verso di esse, ma non se ne conosce bene il significato. In definitiva, le cellule sensoriali esterne servono principalmente per amplificare il movimento della membrana tettoria, che, a questo punto, stimola le ciglia delle cellule sensoriali interne. Quest’ultime sono le vere cellule recettrici dello stimolo, che viene poi passato alle fibre del sistema nervoso dirette verso il cervello. |
E’ evidente che in una determinata specie le capacità di percepire alcune frequenze e non altre sono da ricercare nella caratteristiche strutturali dell’organo del Corti, che gioca un ruolo decisivo nelle capacità percettive delle diverse specie dei vertebrati.
L’organo del Corti non è solo un sistema in grado di discriminare le frequenze e le intensità dei suoni, ma ha anche altre funzioni. In presenza di suoni di forte intensità può sopprimere tutti quelli meno forti emessi contemporaneamente. Questo è molto importante perché consente di eliminare rumori di fondo che potrebbero disturbare la percezione di un segnale biologicamente significativo. Il fenomeno si spiega perché un impulso più forte provoca una sorta d’insensibilità temporanea delle cellule sensoriali esterne contrattili, intorno al punto della membrana basilare in cui è recepito il segnale più intenso. In questo modo sono specialmente eliminati i suoni di frequenze vicine, ma di minore intensità.
Un altro interessante aspetto della percezione acustica è l’accentuazione della risposta all’inizio di un suono per favorire la ricezione di uno stimolo iniziale. L’effetto è legato in questo caso alla capacità di modulazione delle fibre sensoriali connesse alle cellule recettive interne.
Come per la frequenza anche la sensibilità verso l’intensità dei suoni è diversa tra le specie. Anche questo è da collegare alla diversa struttura dell’organo del Corti, ma anche alle capacità amplificative delle strutture auricolari. Molti animali percepiscono suoni di debolissima intensità, con sensibilità molto superiori a quelle dell’uomo.
Si è osservato che le capacità percettive dei suoni si abbassano con l’età e ciò può avere diverse cause. Oltre ai fenomeni di sordità legati alle disfunzioni dell’orecchio medio, di cui si è già accennato, c’è anche il danneggiamento progressivo dell’organo del Corti. In particolare sono colpite le cellule ciliate e il danno è irreversibile. Quest’effetto è tanto più rapido quanto più ci si espone continuativamente a rumori forti. E’ singolare osservare come molti cantanti danneggiano il loro stesso udito perché le loro orecchie sono prossime alla sorgente, che in alcuni casi raggiunge anche 140 dB. In più, uno studio su popolazioni di beduini del deserto giordano ha evidenziato che gli anziani avevano una capacità percettiva equivalente a quella dei giovani perché cresciuti in un ambiente scarsamente rumoroso. |