Struttura del capello e dei follicoli piliferi
Struttura del capello e dei follicoli piliferi
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Le informazioni di medicina e salute contenute nel sito sono di natura generale ed a scopo puramente divulgativo e per questo motivo non possono sostituire in alcun caso il consiglio di un medico (ovvero un soggetto abilitato legalmente alla professione).
Struttura del capello e dei follicoli piliferi
Struttura del capello e dei follicoli piliferi
La molecola fondamentale che costituisce il pelo è la cheratina che per le sue caratteristiche di compattezza, resistenza, elasticità e durezza viene definita come "cheratina dura" (sclerocheratina).
La cheratina del pelo è composta da 18 aminoacidi. All'analisi cromatografica di peli normali si trovano in abbondanza cistina, cisteina, serina, glutammina, arginina, asparagina, prolina, glicina, valina, leucina, isoleucina, e in quantità minori altri aminoacidi come triptofano, treonina, tirosina, fenilalanina, arginina, alanina, metionina.
AMINOACIDI individuati nella cheratina dura del capello (in %).
aminoacidi con catena laterale di tipo idrocarburo:
1 glicina 4,1 - 4,2
2 alanina 2,8
3 valina 5,5 - 5,9
4 leucina 6,4 - 8,3
5 isoleucina 4,7 - 4,8
6 fenilalanina 4,1 - 4,2
7 prolina 4,3 - 9,6
aminoacidi con catena laterale di tipo idrossilico:
serina 7,4 - 10,6
9 treonina 7,1 - 8,5
10 tirosina 2,2 - 3,0
aminoacidi con catena laterale di tipo acido:
11 acido aspartico 3,9 - 7,7
12 acido glutammico 13,6 - 14,2
aminoacidi con catena laterale di tipo basico:
13 arginina 8,9 - 10,8
14 lisina 1,9 - 3,1
15 istidina 0,6 - 1,2
aminoacidi con catena laterale di tipo eterociclico:
16 triptofano 0,3 - 1,3
aminoacidi con catena laterale con contenuto di zolfo:
17 cistina 16,6 - 18,0
18 metionina 0,7 - 1,0
In effetti la molecola di cheratina, assai complessa, è essenzialmente costituita da due proteine assai diverse tra loro sia per il contenuto aminoacidico che per la organizzazione molecolare.
Uno dei due costituenti proteici della cheratina è formato da un filamento non molto ricco di aminoacidi solforati, la cheratina filamentosa, a sua volta costituita da una quota lineare ed una organizzata ad alfa elica in cui si trovano gli aminoacidi solforati (soprattutto cisteina ).
L'altro costituente proteico della cheratina è invece molto ricco di aminoacidi solforati, non ha forma filamentosa, e viene definito come matrice amorfa della cheratina.
A livello dell'epitelio germinativo del bulbo pilifero la matrice fondamentale si dovrà trasformare nei tre diversi componenti del fusto del pelo: cuticola, corteccia, midollo.
La cuticola è costituita da una sola filiera di lunghe cellule cheratinizzate, prive di pigmento e di forma quadrangolare che, embricandosi con il bordo libero rivolto verso l'alto, formano il tipico disegno "a tegolato" della superficie del capello. Per sovrapposizione appaiono, in sezione, come formare 4 -7 strati.

La corteccia o "corticale"è costituita da cellule pigmentate, ricche di melanina, che formano i cosiddetti "fusi" o "cellule fusiformi". I fusi si formano per tappe successive. Inizialmente si legano "testa-coda" le catene proteiche filamentose di cheratina.

Successivamente, come seconda tappa, si formano ponti disolfuro, tra le molecole di cisteina (che diventa cistina) del filamento cheratinico, che porteranno al formarsi della tipica alfa elica della cheratina (cheratina alfa).
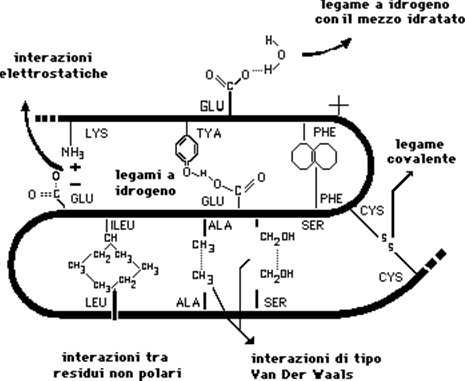
La terza tappa si realizza per il legame tra filamenti diversi di cheratina alfa tramite ponti disolfuro fra molecole di cistina. Si formano così lunghe catene di alfa cheratina legate insieme due o tre alla volta a costituire il "protofilamento" o "protofibrilla".
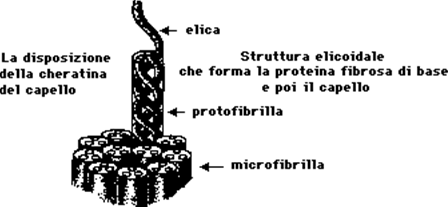
Alla quarta tappa più protofilamenti (sei o sette) vengono cementati fra di loro dalla matrice amorfa proteica, ricca di aminoacidi solforati, sempre con ponti disolfuro, a costituire l'entità elementare della struttura cheratinica: il "tonofilamento" o "microfibrilla" (circa 85 A° di diametro). Infine, un grande numero di tonofilamenti, sempre uniti fra di loro da ponti disolfuro, costituiscono un "fuso" o "cellula fusiforme della corticale"; molti fusi formeranno infine la corticale del capello. Tra un fuso e l'altro si trovano granuli di pigmento melanico e bollicine d'aria; la densità del pigmento e la quantità delle bollicine determineranno il colore del capello. Il processo di cheratinizzazione si completa solo a livello del colletto del pelo.
Il midollo, che è quasi assente sopra la zona della matrice, ben rappresentato in prossimità del bulbo e tende poi a diminuire distalmente nel fusto, è costituito da grandi cellule cheratinizzate, lassamente connesse, disposte in filiere ordinate contenenti granuli di melanina, "bollicine" d 'aria e filamenti di cheratina.
Il pelo è quindi una struttura cheratinica di forma cilindrica, con diametro di circa 65-78 micron (1 micron = 1 millesimo di millimetro) nella razza latina, derivante da una invaginazione epidermica detta follicolo pilifero, appositamente specializzata. Il follicolo pilifero forma con il piano cutaneo un angolo acuto di circa 75°.
La struttura completa del follicolo pilifero consta di due porzioni, una superiore stabile, ed una inferiore caduca; il punto di confine fra queste due parti è detto colletto del pelo.
Alla base di questa complessa struttura troviamo un piccolo bottone dermico che la compenetra: la papilla. La papilla dermica, con la sua ricca vascolarizzazione, nutre uno strato di particolari epiteliociti (tricocheratinociti) che sono con questa in stretto contatto e dai quali prende origine il pelo; questa struttura epiteliocitica è detta matrice del pelo. Le cellule della matrice, moltiplicandosi, si sospingono l'un l'altra verso l'alto all'interno del follicolo e, cheratinizzandosi (zona cheratogena del bulbo), danno luogo alla formazione del pelo. La moltiplicazione cellulare dei tricocheratinociti e la sintesi della cheratina avvengono sotto la spinta di un "messaggio autocrino", presumibilmente un fattore di crescita della famiglia dell'EGF che potremmo definire come Hair Growth Factor (HrGF), e sono più attive nelle zone periferiche della matrice, forse perché più lontane da un "messaggio paracrino inibitorio" prodotto dalla papilla dermica (verosimilmente il TGF beta); così il pelo assume la sua caratteristica forma a tubo e si crea la zona midollare.
Matrice del pelo e zona cheratogena costituiscono il bulbo. Nel pelo normale, fra la matrice e la zona cheratogena, si nota, più o meno accentuata, una strozzatura del bulbo stesso che assume un aspetto leggermente a clessidra. I melanociti, presenti subito al di sopra delle cellule germinative della matrice, cedono pigmento melanico alle cellule in via di cheratinizzazione ed il pelo prenderà il suo caratteristico colore via via che avanza lungo il follicolo e via via che i cheratinociti che lo compongono si trasformano in cellule cornee.
Il pelo, come già accennato, ha tre strati: la cuticola, la corteccia ed il midollo. Il follicolo ha una guaina connettivale, una guaina epiteliale esterna (guaina propria del follicolo) ed una guaina epiteliale interna (guaina propria del pelo) che, presente dalla matrice al colletto, si trova all'esterno della cuticola del pelo e che, al colletto, precisamente a livello dello sbocco della ghiandola sebacea, scompare con desquamazione delle sue cellule cheratinizzate che si mescolano col sebo. Annessa alla porzione superiore e stabile del follicolo vi è una ghiandola che produce un secreto grasso, detto sebo, e lubrifica il pelo aprendosi nel follicolo stesso poco sopra del colletto (ghiandola sebacea).
Attaccato al derma papillare ed a livello della parte inferiore del terzo medio del follicolo vi è anche un piccolo muscolo: il muscolo erettore del pelo, dotato di fibre motorie, si contrae sotto stimolo adrenergico ed è responsabile del fenomeno dell'orripilazione (la "pelle d'oca").
Ogni follicolo pilifero è dotato di un ricco plesso nervoso nel quale sono state ben identificate fibre sensitive (Montagna W.). L'alta capacità percettiva della pelle umana è in gran parte correlata con l'innervazione dei suoi follicoli piliferi e l'uomo è l'unico mammifero i cui follicoli piliferi sono tutti dotati di capacità tattile e dolorifica (capacità che negli altri mammiferi è tipica solo delle vibrisse). Parte cospicua del plesso nervoso del follicolo pilifero è però costituita da fibre simpatiche noradrenergiche ed è stato dimostrato che la simpacectomia provoca una più rapida crescita dei peli (Balus I.) attribuibile all'eliminazione di un effetto inibitorio o all'aumento del flusso ematico locale (Ferguson K. A.).
I follicoli piliferi sono distribuiti su tutta la superficie cutanea, mancano solo sulle palme delle mani, sulle piante dei piedi, sulle falangi distali, sulla cute del pene e sulle semimucose. I peli che ne derivano sono fondamentalmente di due tipi:
-peli piccoli e quasi invisibili, lanugine o vellus, sugli orecchi, sulla fronte, sul tronco e sulle guance delle donne
-peli grossi e pigmentati, peli terminali, presenti solo in alcune sedi e con caratteristiche diverse da zona a zona: lunghi e morbidi sul cuoio capelluto (capelli), corti ed irregolari alle ascelle ed al pube, corti e rigidi alle palpebre (ciglia) e sulle arcate orbitarie (sopracciglia), discretamente lunghi e grossi alle guance maschili (barba).
Il CAPELLO, che fa parte dei peli terminali, si trova in un particolare alloggio della pelle, a forma di sacco, inclinato di circa 75° rispetto alla linea della superficie cutanea, denominato FOLLICOLO PILIFERO. La parte inferiore del follicolo pilifero si trova normalmente nella parte più profonda del derma ma il livello è variabile da capello a capello (la profondità media è di circa 0,6-1 cm). In corrispondenza del terzo superiore del follicolo pilifero sbocca la ghiandola sebacea, per cui l'insieme viene più propriamente definito follicolo pilosebaceo. Al di sotto della ghiandola sebacea, sulla parete esterna del follicolo, è ancorata l'estremità inferiore del muscolo erettore del pelo. Sia il muscolo che la ghiandola sono sempre posti dalla parte dell'angolo ottuso formato dall'asse del follicolo con la superficie orizzontale dell'epidermide (in questo modo la contrazione del muscolo fa assumere al pelo una posizione verticale).
Nel follicolo, dalla superficie scendendo in profondità si distinguono:
a) uno sbocco sulla superficie cutanea, OSTIO, da non confondere con il poro sudoriparo (sbocco della ghiandola sudoripara eccrina);
b) una zona compresa fra ostio e colletto, INFUNDIBOLO, dove fra pelo e parete del follicolo è presente uno spazio libero, normalmente riempito di aria, sebo e detriti cellulari;
c) una strozzatura a livello del terzo superiore, COLLETTO, nel cui punto inferiore sbocca la ghiandola sebacea e termina, desquamando, la guaina epiteliale interna;
d) una parte intermedia o ISTMO, fra colletto e attacco inferiore del muscolo erettore del pelo;
e) infine, al di sotto dell'istmo stesso, una PARTE PROFONDA, alla cui base, cioè alla base del follicolo stesso, è presente un'incavatura rivolta verso l'alto, delimitata dalle cellule della matrice e contenente una struttura connettivale, una specie di "gomitolo" di vasi sanguigni, linfatici e strutture nervose, la PAPILLA, che ha il compito di provvedere al nutrimento ed al controllo endocrino delle cellule della matrice del pelo.
Sempre nel follicolo, dall'esterno verso l'interno, si riconoscono:
1) la GUAINA CONNETTIVALE, che è la più esterna, ricca di terminazioni nervose e formata da strati concentrici e longitudinali di fibre collagene con numerose cellule (fibroblasti);
2) la MEMBRANA VITREA, diretta continuazione della membrana basale cutanea (rispetto alla quale ha perduto il caratteristico aspetto ondulato);
3) la GUAINA EPITELIALE ESTERNA, diretta continuazione degli strati più profondi dell'epidermide che si approfondano seguendo la membrana vitrea, le cellule che la compongono, in basso, in corrispondenza della papilla, assumono le caratteristiche proprie della matrice del pelo; la guaina epiteliale esterna presenta dal lato, quello in cui il follicolo forma con il piano cutaneo un'angolo ottuso, un'apertura irregolarmente circolare attraverso cui passa il dotto escretore della ghiandola sebacea; più in basso questa guaina diventa più spessa formando una sporgenza, sulla quale si trova il punto inferiore di ancoraggio del muscolo erettore del pelo;
4) la GUAINA EPITELIALE INTERNA, formata da tre strati di cellule che originano, come quelle del pelo, dalle cellule della matrice; il suo compito è quello di ancorare il capello al follicolo e formare un involucro più rigido rispetto al pelo che contiene in modo da modellarlo ed orientarlo correttamente. La guaina epiteliale interna, che è quindi a diretto contatto con il pelo, termina in corrispondenza del colletto dove le sue cellule desquamano all'interno dell'infundibolo e si mescolano al sebo.
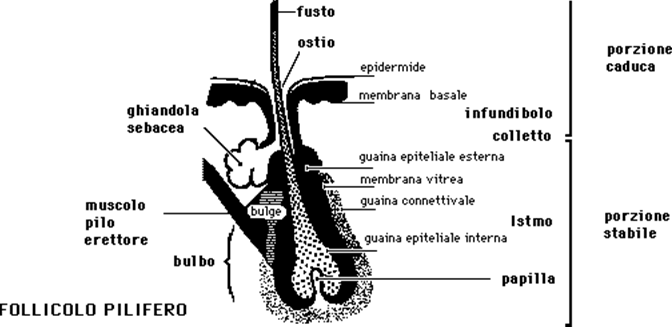
Nel capello (o nel pelo) normalmente sviluppato si distinguono tre parti:
I°) una esterna al follicolo, visibile, STELO o FUSTO. Ha uno spessore nel capello dell'adulto medio di 65-78 micron;
II°) una interna al follicolo stesso, fra l'ostio del follicolo e l'attacco inferiore del muscolo erettore del pelo, RADICE, "immersa" nella cute e quindi normalmente non visibile;
III°) una ancora sottostante, alloggiata nella porzione più profonda del follicolo, BULBO, caratterizzata alla sua base, da due - tre file di cellule sovrapposte a rapida riproduzione che costituiscono la MATRICE.
Quelle della matrice sono le sole cellule capaci di mitosi del pelo e, pertanto, tagliare i capelli corti non può "rinforzarli" in quanto quello che si taglia è una parte di fusto, "un polimero di cheratina" formato cioè da cellule ormai prive di attività vitale (Il solo vantaggio che si può ottenere tagliando corti i capelli è semmai dovuto alla minore traumatizzazione con lavaggi, spazzolature etc. ed alla maggiore facilità di impiego di farmaci in forma di lozioni o shampoo); il capello non deve essere paragonato ad una pianta che può trarre beneficio da "potature" periodiche.
Le cellule della matrice, via via che si riproducono, spingono verso l'alto quelle più vecchie; durante la risalita queste cellule, in modo analogo a quelle dell'epidermide, elaborano nel loro interno la cheratina, vanno cioè incontro al cosiddetto "processo di cheratinizzazione", diventando progressivamente sempre più rigide. Di queste cellule, all'inizio tutte uguali, alcune che vanno a costituire la guaina epiteliale interna si cheratinizzano ed induriscono in anticipo rispetto al resto del pelo (cuticola, corteccia e midollo) e così il pelo mentre cresce si modella nella guaina come in uno stampo ed assume la sua forma definitiva. Subito al di sopra delle cellule delle matrice sono presenti alcuni melanociti che hanno il compito di "colorare", immettendovi la melanina, le cellule che diventeranno la "corteccia" del capello.
Le vere cellule germinative staminali del capello non sono quelle della matrice ma si trovano nella così detta "zona protuberante" (bulge) più alta nel follicolo, a livello dell'istmo (Cotsarelis G.).
Quando si arriva alla fase catagen la matrice, comunemente intesa, degenera e la papilla rimane unita al bulbo solo mediante una specie di "sacco", formato dalla guaina epiteliale esterna che contiene le ultime cellule prodotte dalla attività mitotica sotto forma di una lunga colonna di cellule epiteliali.
Questo sacco di cellule epiteliali poi si deconnette dalla papilla e risale fino all'istmo, in qualche modo attivando la produzione di HrGF delle cellule germinative della zona protuberante. Queste ultime, con un processo molto simile a quello che si osserva nella formazione embriologica del pelo primitivo, entrano in rapida mitosi e migrano verso il basso colonizzando nuovamente la zona della matrice e dando inizio al nuovo anagen; la sede nel bulge delle cellule staminali potrebbe spiegare i molti insuccessi nel tentativo di ottenere in laboratorio delle colture cellulari di tricocheratinociti capaci di dare origine ad un pelo: sarebbero state infatti sempre utilizzate le cellule della matrice, cioè cellule sbagliate!
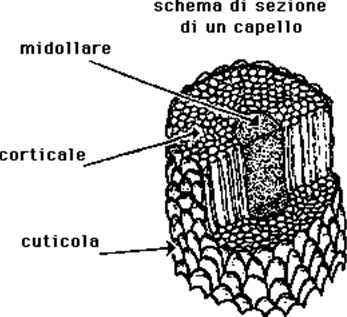
Sezionando un capello in senso orizzontale se ne può vedere la struttura, suddivisibile in tre parti:
a) CUTICOLA, è la parte esterna del capello apparentemente formata, al microscopio a scansione, da 4 -7 filiere di cellule trasparenti e sottili, disposte verticalmente a livello della radice ed invece, a livello del fusto, disposte obliquamente, a "scaglie" embricate, con l'estremità inferiore attaccata alla corteccia e quella superiore staccata, con aspetto quindi a spiga di grano o forasacco.
All'interno del follicolo la cuticola del pelo nasce, in realtà, come un unico strato cellulare. Le cellule divengono cuboidali e quindi piatte via via che salgono nel follicolo e contemporaneamente si aprono verso l'alto e, raggiunto lo stato di completa cheratinizzazione, si sovrappongono come le tegole di un tetto (embricazione).
Nel pelo definitivo le sezioni mostrano la cuticola come una struttura pluristratificata in media composta di sette strati all'estremità inferiore della radice. E' tuttavia, in realtà un solo strato di cellule allungate ciascuna approssimativamente lunga 40 - 50 micron e spessa da 0,5 a 10 micron con molteplici sovrapposizioni e con il margine libero delle cellule superficiali aperto verso l'alto. Si ha così un aspetto microscopico pluristratificato simile ad una pasta sfoglia.
Data la posizione in cui si trova, la cuticola è la prima ad essere danneggiata quando il capello viene "maltrattato" con shampoo inadeguati, permanenti, spazzolature eccessive etc;
b) CORTECCIA, intermedia, è la parte più abbondante; è formata da cellule più grosse, di forma fusata, lunghe 90 micron e larghe 5 micron, disposte verticalmente in file parallele; contengono un pigmento, la melanina, che tende a ridursi con l'età facendo diventare il capello "bianco" (incanutimento). Da quanto detto risulta chiaro che per "decolorare" il capello, ad esempio con acqua ossigenata, è necessario che la reazione chimica avvenga a livello della corteccia dato che la cuticola, più esterna, non è colorata, così questa, se il capello viene esaminato al microscopio a forte ingrandimento dopo una decolorazione malfatta può presentarsi gravemente danneggiata o addirittura scomparsa;
c) MIDOLLO, interno, formato da cellule arrotondate, disposte a colonne, generalmente separate fra loro da spazi d'aria (l'aria trattenuta ha funzione determinante per la protezione dal freddo e per questo motivo il midollo supera negli animali il 50% dello spessore totale del pelo mentre è scarsamente rappresentato e talora assente nella specie umana).
Ghiandola sebacea
E' posta lateralmente al follicolo pilifero e ha il compito di produrre il SEBO, sostanza grassa acida a pH medio 3,5, orientativamente così composta (sulla superficie cutanea): trigliceridi 30-42%, cere 20-25%, acidi grassi liberi 15-25%, squalene 9-12%, idrocarburi ramificati 4-8%, colesterolo esterificato 2-3%, colesterolo libero 1-1,5%, altre sostanze come di e monogliceridi, glicerolo etc. 3-5%.
Il sebo ha due funzioni principali:
la prima è di contribuire, con il sudore, alla formazione del film idro-lipidico di superficie, una emulsione acqua/olio che protegge la superficie cutanea dalle aggressioni chimiche (detergenti, solventi etc.) e batteriche; la seconda è di lubrificare e impermeabilizzare la superficie esterna del capello via via che questo allunga.
Il dotto escretore della ghiandola sebacea si apre nella parte superiore del follicolo in modo da lubrificare il capello prima ancora che questo compaia alla superficie cutanea. Sul cuoio capelluto la produzione totale di sebo è di 650 - 700 mg nelle 24 ore. Il film idrolipidico (emulsione acqua/olio formata dall'unione del sebo con l'acqua del sudore) dopo asportazione con detergente si ricostituisce in un tempo relativamente breve (3 - 6 ore) ma la ghiandola sebacea non sembra, al contrario di quanto spesso affermato, poter aumentare la propria attività rispetto ai valori basali per lavaggi frequenti.
Il sebo non ha la possibilità di spostarsi in modo autonomo lungo il fusto del capello. Il fatto che i capelli possono rapidamente e nuovamente "ungersi" dopo un lavaggio è dovuto alla riduzione della tensione superficiale fra le "tegole cheratiniche" della cuticola associata al continuo sfregamento fra i capelli. La velocità di migrazione del sebo aumenta se i capelli vengono continuamente toccati con le mani o pettinati. Inoltre, nei soggetti con capelli particolarmente e rapidamente untuosi, il sebo è più fluido e quindi ha più alta capacità di migrazione.
Nei soggetti con seborrea (vedi) l'abbondanza di sebo provoca inconvenienti estetici dovuti alla facile adesione delle polveri ambientali, dei residui cheratinici, delle sostanze di rifiuto veicolate dal sudore. Il tutto, troppo spesso, accentuato dall'impiastricciamento dei cosmetici applicati.
La secrezione delle ghiandole sebacee è controllata dagli ormoni steroidi, circolanti o prodotti localmente dal follicolo pilosebaceo a partire dai precursori di origine gonadica e surrenalica (il più attivo in tal senso sembra essere l'androstandiolo).
Le ghiandole sebacee producono abbondantemente sebo durante la vita fetale (vernice caseosa del feto). Sono attive nei primi mesi di vita. Passano in una fase di relativa quiescenza fino ai 9-10 anni di età per riattivarsi all'adrenarca. Ritrovano poi piena attività alla pubertà e sono causa di fenomeni di piccola patologia dermatologia, di cui è prototipo il fenomeno dell'acne polimorfa comune giovanile. La produzione di sebo subisce una brusca riduzione nella donna alla menopausa mentre nel maschio, ed in modo assai personale, diminuisce solo molto lentamente dopo i 60 - 70 anni di età.
Muscolo pilo-erettore
E' disposto obliquamente sullo stesso lato della ghiandola sebacea, dalla parte dell'angolo ottuso formato dall'asse del follicolo con la superficie orizzontale dell'epidermide.
Delle due inserzioni, quella profonda è sulla guaina epiteliale esterna, a livello dell'istmo, sulla parte inferiore del terzo medio del follicolo, (zona protuberante o bulge) mentre quella superficiale si ancora al derma papillare con un sistema di fibre dissociate su un'area relativamente vasta.
Il BULGE non è una singola protuberanza (Misciali C.) ma è costituito da multiple formazioni digitiformi che prendono origine da una porzione della circonferenza dell'istmo, ognuna di queste appendici è connessa ad un fascio del muscolo erettore del pelo.
La funzione del muscolo erettore è di raddrizzare il pelo, contraendosi ("orripilazione"): l'aria che rimane intrappolata fra i peli si riscalda a contatto con la superficie corporea ed essendo una cattiva conduttrice di calore protegge la cute dal freddo ambientale.
Secondo alcuni Autori la contrazione del muscolo pilo-erettore potrebbe favorire, per compressione, lo svuotamento della ghiandola sebacea. Il muscolo erettore è assente nei peli di tipo lanuginoso.
Costituzione chimica del capello
All'analisi chimica i costituenti principali del capello, oltre all'acqua, sono: cheratina, lipidi, minerali e pigmenti.
Cheratina: è una proteina contenuta principalmente nella corteccia (della quale ne costituisce la massima parte); gli aminoacidi presenti in maggiore quantità nella cheratina sono la cisteina e la cistina (derivata dalla condensazione di due molecole della prima con perdita di acqua e formazione di legami disolfuro), la serina, l'acido glutammico, la glicina, la tretionina, l'arginina, la valina, la leucina e l'isoleucina. La cheratina presente in maggiore quantità nel capello è quella alfa, fibrosa, a basso contenuto di zolfo, con Peso Molecolare di circa 45.000, insolubile in acqua.
Le catene polipeptidiche sono rese stabili da tre tipi di "ponti": legami idrogenati (conferiscono solidità), ponti fra catene acide e catene basiche (si rompono con gli acidi forti) e ponti disolfurici (quando sono lesi, ad esempio nelle "permanenti", il capello si arriccia). La cheratina può essere deformata con il vapore acqueo ("messa in piega").
Il processo di cheratinizzazione, in particolare quello dei peli, è regolato da ormoni, vitamine, fattori genetici e metabolici e sembra legato al metabolismo del colesterolo e alla sua esterificazione con acidi grassi sintetizzati dall'epidermide. Ne consegue che carenze dietetiche e/o difetti enzimatici delle vie di sintesi del colesterolo e degli acidi grassi possono provocare una cheratinizzazione anomala con conseguenti difetti strutturali del fusto.
Lipidi: costituiti da trigliceridi, cere, fosfolipidi, colesterolo, squalene ed acidi grassi liberi sono quantitativamente documentabili con estrema difficoltà in quanto in massima parte derivati da quelli del sebo.
Minerali (oligoelementi): rappresentano una componente essenziale dei sistemi proteico-enzimatici. Esiste una correlazione diretta fra la quantità di oligoelementi presenti nel sangue e quelli presenti nel capello.
a) ferro (media 4-12 mg/gr di capelli): è più abbondante nei capelli rossi rispetto a quelli biondi e neri.
b) magnesio (media 30-45 mg/gr): è più abbondante nei capelli di colore nero (fino a 170 mg/gr). Catalizza la trasformazione dell'ATP in cAMP, in seguito alla attivazione della adenilciclasi, e la degradazione dell'cAMP in 5-AMP ad opera della fosfodiesterasi (vedi).
c) zinco (media 150-180 mg/gr): è indispensabile per la corretta funzione dei fattori di crescita e quindi per la attività delle cellule germinative della matrice. In sua carenza il capello si indebolisce e rallenta il ritmo di crescita.
L'eccesso di zinco determina carenza di rame a causa dell'induzione dell'enzima tioneurina che, a livello intestinale, lega il rame in modo preferenziale rallentandone o impedendone l'assorbimento.
d) rame (media 16-50 mg/gr): è indispensabile per catalizzare la conversione della tirosina a DOPA (nel processo di sintesi della melanina) e per consentire l'ossidazione della cisteina in cistina con formazione dei ponti disolfuro.
e) piombo (media 10-30 mg/gr): è più abbondante nei capelli di colore castano. Dato che la quasi totalità del piombo presente nell'organismo deriva da quello esogeno e poiché il capello ne rappresenta la sede principale di accumulo, la sua misurazione a livello della parte distale del fusto viene utilizzata per valutare la presenza di questo metallo nell'inquinamento ambientale.
La carenza di proteine e/o sali minerali potrà essere evidenziata, al microscopio, da un fusto sottile associato a bulbi molto piccoli (se il capello è costituzionalmente sottile i bulbi appariranno invece normali).
Pigmenti: sono rappresentati dalle melanine (sostanze colorate), presenti nella corticale del pelo in forma diffusa o granulare. Sono insolubili in acqua, solubili negli acidi forti, decolorabili con acqua ossigenata.
I melanociti, utilizzando la tirosina come precursore, sintetizzano due principali tipi di melanina: l'eumelanina, scura e presente nei capelli neri e la feomelanina, più chiara e presente nei capelli dorati, biondi o rossi.
BIBLIOGRAFIA
Auber L.: "The anatomy of follicles producing wool fibres with special reference to keratinization" Trans Roy soc Edin 1952; 62: 19.
Balus I.: "Experiments on the relationship between the nervus system and hair" Brit J Dermatol 1961; 73: 69.
Bosco I.: "Dermatologia generale" Roma, SEU, 1970. 32-35.
Caputo R., Alessi E.: "Istologia della cute e degli annessi cutanei" in: Serri F. "Trattato di dermatologia" PICCIN, Padova, 1986,1°. 38-46 .
Duni D., Cislaghi E.: "Elementi di tricologia" Milano, SEPeM, 1988. 59-62.
Enjolras O.: "Centomila capelli" Parigi, Parente L.,1978. l0-11.
Ferguson K. A.: "The effect of sympacectomy on wool growth" Aust J Sci Res 1949; 2: 438.
Frati C., Didona B.: "I fattori di crescita e le loro implicazioni in dermatologia" Chron Derm 1991; 3 : 395.
Misciali C.: "Studio istologico dell'istmo del follicolo pilifero" Abstracts 70° Congresso Nazionale della Società Italiana di Dermatologia e Venereologia, Editrice C.S.H., Milano, 1995, 420.
Montagna W.: "The structure and function of skin" 2nd ed., Acad Press, New York, 1962, 174 - 267.
Montagna W.: "Morfology of cutaneous sensory receptor" J Invest Dermatol 1977; 69: 4.
Paus R., Heinzelmann T., Schultz K.D., et al: "Hair growth induction by substance P" Lab Invest 1994; 71: 134 -140.
Sturaro A., Parvoli G., Doretti L., et al: "The influence of color, age, and sex on content of zinc, copper, nikel, manganese, and lead in human hair" Biol Trace Elem Res 1994; 40: 1 - 8.
Fonte: estratto / citazione da http://www.sitri.it/marliani.doc
"TRICOLOGIA" -diagnostica e terapia- II edizione elettronica 1997
visitate il sito : http://www.sitri.it/
Autore del testo:
ANDREA MARLIANI
Proprietà letteraria ed artistica riservata all'Autore.©
Parola chiave google : Anomalie dei capelli tipo file : doc
Struttura del capello e dei follicoli piliferi
Visita la nostra pagina principale
Struttura del capello e dei follicoli piliferi
Termini d' uso e privacy