Cervello umano
Cervello umano
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Cervello umano
IL CERVELLO E LE SUE FUNZIONI
Introduzione
In questo saggio si cerca di spiegare in modo generale il funzionamento del cervello. Argomenti della trattazione sono: la percezione, la memoria, il movimento volontario ed involontario.(parleremo del linguaggio in un saggio successivo)
La trattazione di questi argomenti è fatta seguendo pochi principi di carattere generale, tali che l’intero sistema cerebrale risulta obbedire a poche regole applicabili anche a sistemi cerebrali di animali inferiori.
Nel testo vi sono numerosi esempi e ripetizioni di concetti. Ho preferito la chiarezza ad un testo formalmente ineccepibile.
La nostra trattazione segue un principio generale che è il seguente
La mente o il cervello ha sempre una funzione attiva e costruttiva.
Tutte le patologie devono essere spiegate come un difetto di costruzione. Possiamo immaginare il cervello come una fabbrica che produce oggetti complessi come le automobili. Essa è suddivisa in svariati settori nei quali sono prodotti, assemblati e verniciati i molteplici componenti che determinano il prodotto finale, in questo caso l’automobile. Se un settore non funziona bene, le macchine sono difettose.
Ciò che appare incredibile è il fatto che per la costruzione delle automobili occorrono giorni, per la costruzione del mondo esterno il cervello impiega frazioni di secondo. Microsecondo per microsecondo edifichiamo e riedifichiamo non solo tutto ciò che appare esterno a noi stessi, ma anche i nostri pensieri, ricordi, emozioni…
Questo principio generale è importante poiché in modo più o meno consapevole i neuroscienziati sono “realisti”. Essi sono convinti che esista una “realtà esterna” a noi. Essa è composta da “oggetti”, con la loro forma, struttura, dimensione, colore; con le loro relazioni spaziali e temporali. Compito del cervello è semplicemente quello di “recepire”, ossia “trasferire” all’interno della mente questa “realtà” preformata.
Le molteplici teorie proposte, partendo da questo presupposto errato, sono palesemente infondate, alcune addirittura assurde.
LA PERCEZIONE
Le conoscenze attuali
Tutti i sistemi sensoriali si basano su principi generali comuni, ma, essendo quello visivo il più studiato, tratteremo soprattutto della percezione visiva.
Si ritiene che la visione comporti l’intervento di tre vie poste in parallelo, che elaborano separatamente le informazioni relative al movimento, alle forme ed al senso della profondità, e ai colori. Si tratta dei sistemi
magnocellulare (movimento);
parvicellulare interblob (forme e senso della profondità)
parvicellulare blob (colori)
Il sistema magnocellulare è specializzato per l’analisi del movimento e delle relazioni spaziali degli oggetti; esso contribuisce anche alla visione stereoscopica. Questa via prende inizio dalle grandi cellule gangliari di tipo M della retina, che proiettano agli starti mognocellulari del corpo genicolato laterale del Talamo. La via continua nello strato IVC e quindi negli strati IVB e VI di V1. Da qui essa arriva alle strisce spesse di V2, prosegue in V3 e da qui in MT (V5), che è l’area che Zeki ha osservato essere connessa con l’analisi del movimento e del senso della profondità. I neuroni di questo sistema forniscono a tutti i livelli, risposte rapide e transitorie. Sono relativamente insensibili ai colori. E’ stato osservato che il sistema magnocellulare ha una certa capacità di contribuire alla percezione del senso di profondità, mentre è del tutto inefficiente per l’analisi degli oggetti statici.
“Il sistema parvo proietta allo strato Cbeta dell’area 17 e da qui a due raggruppamenti cellulari degli strati 2 e 3 della stessa area, che si colorano diversamente con la citocromo ossidasi: quelli scuri sono chiamati blob (bolla), quelli chiari interblob
Le cellule blob sono sensibili al colore e proiettano alle striscie sottili di V2 e da qui all’area V4 (giro fusiforme). Le cellule interblob si connettono con le strisce pallide di V2 che sono sensibili all’orientamento delle linee e sono connesse con V3.
In conclusione, movimento e stereopsi sono competenza prevalentemente del sistema magno e hanno stazione terminale in V5. Il colore è analizzato dal sistema parvo ed ha stazione terminale in V4. Cellule sensibili alla direzione e quindi impegnate nella percezione della forma sono presenti in più aree ma particolarmente in quelle V2, V3 e V4. Le ulteriori proiezioni di questi sistemi seguono probabilmente due grandi vie, ventrale e dorsale, che, secondo Mishkin e coll (1983) trasmettono le informazioni elaborate dalle aree visive verso i centri superiori. La via ventrale consiste in una serie di connessioni multisinaptiche, che seguono il decorso del fascicolo longitudinale inferiore e congiungono le aree occipitali con le aree temporali inferiori nelle quali avviene l’identificazione visiva dello stimolo. V4 proietta a queste aree. La via dorsale consiste in connessioni multisinaptiche, che seguono la via del fascicolo longitudinale superiore e connettono le aree striate e prestriate con il lobulo parietale inferiore, nel quale avviene la localizzazione dello stimolo. MT proietta alla corteccia parietale (Livingston e Hubel, 1988) …”
Lo spazio ed il tempo come forme pure della percezione
Nell’Estetica Trascendentale, Kant sostiene che due sono le forme pure dell’intuizione sensibile: lo spazio e il tempo. Essi sono i principi della conoscenza a priori degli oggetti.
Lo spazio è la condizione necessaria di tutti i rapporti, in cui gli oggetti sono intuiti come fuori di noi; il tempo è la condizione necessaria dell’intuizione di noi stessi e del nostro stato interno. Lo spazio è la forma dei cinque sensi esterni; il tempo è la forma del senso interno. Per forma Kant intende il modo di funzionare, ossia la condizione alla quale deve sottostare la rappresentazione sensibile di oggetti esterni ed interni. Kant nega che lo spazio e il tempo siano realtà assolute, indipendenti dalla forma delle nostra intuizione sensibile o che essi possano rappresentare le condizioni o le qualità delle cose. L’essere umano coglie le cose spazialmente e temporalmente determinate solo in quanto possiede una sensibilità strutturata in questo modo. Lo spazio e il tempo sono forme del Soggetto non dell’oggetto; essi hanno realtà empirica e idealità
Si noti infine che tra spazio e tempo, una certa supremazia va riconosciuta al secondo:
“Lo spazio, in quanto forma pura di ogni intuizione esterna, è limitato, come condizione a priori, semplicemente ad apparenze esterne. Al contrario, dato che tutte le rappresentazioni -- non importa che abbiano o no come oggetto cose esterne -- appartengono comunque in sé stesse, come determinazioni dell'animo, allo stato interno, mentre questo stato interno cade poi sotto la condizione formale dell'intuizione interna, e quindi del tempo, il tempo allora è una condizione a priori di ogni apparenza in generale, e più precisamente la condizione immediata delle apparenze interne (delle nostre anime) e proprio per questo, indirettamente, anche delle apparenze esterne (Ragione pura, B 60).
Per Kant, gli oggetti al di fuori della nostra percezione sono apparenze.
“Noi abbiamo dunque voluto dire che tutta la nostra intuizione non è altro che la rappresentazione di un'apparenza (Erscheinung); che le cose da noi intuite non sono in sé stesse così come le intuiamo, e che i loro rapporti non sono costituiti in sé così come appaiono a noi; che se noi sopprimiamo il nostro soggetto, o anche soltanto la costituzione soggettiva dei sensi in generale, in tal caso tutta quanta la costituzione e tutti i rapporti degli oggetti nello spazio e nel tempo, anzi persino lo spazio e il tempo, sono destinati a svanire. Tutte queste cose, in quanto apparenze, non possono esistere in sé stesse, ma esistono soltanto in noi. Di che cosa mai possa trattarsi, riguardo agli oggetti in sé stessi, separati da tutta questa recettività della nostra sensibilità, ci rimane perfettamente ignoto. Noi non conosciamo altro che il nostro modo di percepire gli oggetti” (Ragione pura, B 65)
Figure e sfondi
Nel riquadro sottostante si possono vedere due volti di profilo oppure una coppa. Questa duplice visione dipende da ciò che per il cervello è la figura e ciò che per il cervello è lo sfondo.

La separazione della figura dallo sfondo è una delle funzioni fondamentali della percezione. Senza questa separazione il mondo ci apparirebbe come un insieme indiscriminato di forme e colori.
La nostra teoria sulla percezione
Il meccanismo della percezione deve, necessariamente, essere semplice. Non convincono le teorie attuali che ipotizzano complesse elaborazioni a vari stadi. Infatti, quasi tutti gli animali sono in grado di vedere oggetti in movimento, in brevi frazioni di secondo e, se per osservare un oggetto, il cervello si perdesse in complesse elaborazioni, mancherebbe il tempo per catturare la preda o sfuggire ad un predatore.
Riepilogando quanto finora esposto, vi sono tre componenti essenziali per formalizzare una nuova teoria sulla percezione.
- il sistema visivo ( e come vedremo in seguito, anche gli altri sistemi percettivi) utilizza due vie in parallelo per la costruzione dell’immagine
- lo spazio ed il tempo sono forme pure delle sensazioni esterne e del senso interno
- la separazione della figura dallo sfondo è la funzione fondamentale di ogni percezione
A questi tre punti va aggiunta la considerazione di carattere generale concernente la funzione costruttiva del cervello
Affermiamo che:
- la funzione fondamentale di ogni percezione è quella di costruire per ogni emisfero cerebrale una figura ed uno sfondo
- Le due figure sono costruite dal sistema occipito/temporale; i due sfondi sono costruiti dal sistema occipito/parietale.
- L’emisfero destro costruisce una figura mobile su uno sfondo fisso; l’emisfero sinistro costruisce una figura fissa su uno sfondo mobile.
- Lo sfondo fisso costruito dall’emisfero destro costituisce la dimensione spaziale dello “oggetto”, è il sistema di riferimento spaziale; lo sfondo mobile, costruito dall’emisfero sinistro, costituisce la dimensione temporale dello “oggetto”, è il sistema di riferimento temporale
In questo articolo per semplificare il discorso non ci occupiamo dei colori.
Il sistema magnocellulare o “dorsale” non costruisce i dettagli fini. Esso, infatti, è preposto alla costruzione dei due sfondi. Il sistema parvicellulare o “ventrale” costruisce i dettagli fini della figura, costruisce pure i colori, essendo preposto alla costruzione degli oggetti.
Per capire come ciò accade, facciamo un esempio esplicativo.
Immaginiamo di dover tracciare con la penna una linea su di un foglio. Per poterlo fare è necessario che mentre la penna scorre sul foglio, quest’ultimo stia fermo. Infatti, se anch’esso si muovesse assieme alla mano, la linea non potrebbe essere tracciata.
Vi è però un secondo modo di tracciare la linea. Si può tenere ferma la mano e muovere il foglio sottostante.
Non esiste una terza possibilità. Se penna e mano si muovono contemporaneamente oppure stanno ambedue immobili la linea non può essere tracciata.
L’esempio su addotto spiega abbastanza bene come viene costruito lo “oggetto”.
 Nel nostro esempio, la mano con la matita è il sistema ventrale, il foglio è il sistema dorsale.
Nel nostro esempio, la mano con la matita è il sistema ventrale, il foglio è il sistema dorsale.
Le due figure si inseriscono nei rispettivi sfondi

Quando osserviamo qualcosa, per esempio un bicchiere, il sistema oculomotore segue la forma del bicchiere. Esso percorre un itinerario preciso dettato dalla “forma dello “oggetto”. I numerosi segnali che provengono dalla retina, in particolare quelli relativi alle variazioni di luminosità, eccitano specifiche cellule del sistema colonnare della corteccia visiva primaria. Le cellule sensibili alle variazioni di luminosità scaricano seguendo specifici orientamenti di linee che costituiscono il contorno dello “oggetto”. Ricordiamo che i segnali giungono al cervello ordinati dal sistema retinotopico.
Nel tracciare questo profilo i due sistemi (dorsale e ventrale) devono separare i numerosissimi segnali che provengono dai recettori sensoriali. Devono fare una scelta. Porre l’attenzione su alcuni e scartarne altri. Se non vi fosse questa selezione il sistema crollerebbe sotto il peso delle numerose informazioni.
Gli elementi focalizzati dal sistema ventrale compongono la figura; gli elementi focalizzati dal sistema dorsale costituiscono lo sfondo
Mentre il sistema oculomotore segue la”forma del bicchiere” il sistema ventrale traccia questa forma su uno sfondo immobile, costruito dal sistema dorsale. Questa funzione viene espletata nell’emisfero destro.
Contemporaneamente il sistema ventrale traccia la medesima forma rimanendo immobile; in questo caso è la mobilità dello sfondo costruito dal sistema dorsale che consente di disegnare la figura. Questa funzione è espletata nell’emisfero sinistro.
Immaginiamo di dover effettuare il disegno di due oggetti, un bicchiere e una bottiglia.
Dapprima realizziamo il primo disegno spostando la matita e tenendo fermo il foglio. Successivamente stacchiamo la matita dal foglio, quindi eseguiamo il secondo disegno.
In questo caso i due oggetti disegnati occupano un “posto” diverso nello “spazio”
Effettuiamo adesso lo stesso disegno utilizzando il secondo metodo, tenendo ferma la matita e movendo lo sfondo. Effettuato il primo disegno stacchiamo la matita, nel frattempo lo sfondo è andato avanti, adesso effettuiamo il secondo disegno, sempre movendo lo sfondo. In quest’occasione i due oggetti sono disegnati in “successione temporale”, prima l’uno, poi l’altro.
Nel cervello, lo ribadiamo il sistema dorsale espleta la funzione del foglio, il sistema ventrale quello della matita.
Proviamo a tradurre in termini operativi il concetto di percezione visiva di più oggetti nello spazio. Essa consiste nel disegnare nitidamente gli oggetti spostando la matita su uno sfondo immobile.
Proviamo a tradurre in termini operativi il concetto di rappresentazione di più oggetti in successione temporale. Essa consiste nel disegnare nitidamente gli oggetti, tenendo ferma la matita e movendo lo sfondo.
La costruzione dei vari oggetti
Osserviamo la seguente figura.

Il cervello riconosce immediatamente un rettangolo. Affinché ciò sia possibile, il sistema oculomotore deve tracciare il contorno secondo un preciso percorso che potrebbe essere analogo al seguente

Supponiamo che vi dica di guardare dapprima la base e poi l’altezza del rettangolo. Il cervello costruisce con un specifico tracciato oculomotore i due seguenti segmenti.
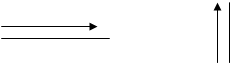
Lo stesso procedimento si può fare con l’angolo, la diagonale, …
L’oggetto che noi vediamo, quindi, altro non è che il risultato di un preciso processo costruttivo del cervello. Tutto dipende da dove viene fissata l’attenzione.
.
Il mondo fisico e il mondo psichico
Consideriamo adesso il mondo fisico. Esso è costituito da oggetti mobili (corteccia occipito/temporale destra) incollati su uno sfondo fisso (corteccia occipito/parietale destra).
Possiamo suddividere lo spazio costituito dal sistema occipito/parietale destro in due parti:
spazio oggettuale. E’ lo spazio del singolo oggetto, della singola figura costruita dal sistema occipito/temporale destro
spazio relazionale. E’ lo spazio di più oggetti posti in relazione l’uno con l’altro
Se sono danneggiati lo spazio oggettuale e lo spazio relazionale si ha difficoltà nel costruire l’immagine dell’oggetto, la sua figura, nonché a ricostruire la posizione nello spazio dei vari oggetti.
Le lesioni alla corteccia parietale destra danneggiano lo spazio oggettuale e lo spazio relazionale.
Consideriamo adesso il mondo psichico. Esso è costituito da oggetti fissi (corteccia occipito/temporale sinistra) incollati su uno sfondo mobile
Possiamo suddividere il “tempo” costituito dal sistema occipito/parietale sinistro in due parti.
tempo oggettuale o durata. E’ il tempo di costruzione del singolo oggetto, della singola figura costruita dal sistema occipito/temporale sinistro
tempo relazionale. E’ il tempo di costruzione di più oggetti o le loro relazioni temporali.
Le lesioni alla corteccia parietale sinistra danneggiano il tempo oggettuale o durata e il tempo relazionale.
IL MOVIMENTO
Affinché si possano percepire oggetti in movimento è necessario che i due sfondi e le due figure si integrino in una percezione unitaria. Il movimento, infatti, comporta la percezione di un oggetto in tempi diversi in luoghi diversi.
Nella percezione visiva del movimento è dominante il sistema occipito/parietale destro e quindi lo spazio; nella esecuzione volontaria o involontaria dei movimenti è dominante il sistema occipito/parietale sinistro e quindi il tempo.

Nella figura la stessa macchina occupa posti diversi (sistema dominante) in successione temporale (sistema sottodominante
Consideriamo adesso il movimento del corpo nello spazio. Supponiamo di muovere la mano destra ad occhi chiusi.
Durante questo gesto si ha una duplice consapevolezza. La prima riguarda la posizione nello spazio della mano. Siamo consapevoli che la mano occupa, durante il movimento posti diversi nello spazio. Ciò dipende dal sistema che costruisce lo “oggetto” mano su uno sfondo fisso (lo spazio). Lo spazio è attivato dal sistema parieto/parietale destro; lo “oggetto” mano è costruito dal sistema parieto/temporale destro.
La seconda consapevolezza riguarda la durata delle contrazioni muscolari che muovono gli arti e i rapporti temporali di queste contrazioni. Siamo consapevoli del tempo che l’arto impiega nel suo movimento.
Ciò dipende dal sistema che costruisce lo “oggetto” mano su uno sfondo mobile (il tempo). Il tempo è attivato dal sistema parieto/parietale sinistro; lo “oggetto” mano è costruito dal sistema parieto/temporale sinistro.
L’integrazione di queste due consapevolezze costituisce la consapevolezza del movimento dell’arto nello spazio.
LE PATOLOGIE DEL SISTEMA OCCIPITO/PARIETALE
La costruzione dei due sistema di riferimento si completa nella corteccia parietale. Solo le patologie della corteccia parietale destra danneggiano solo lo “spazio oggettuale” e lo “spazio relazionale”; analogamente solo le patologie alla corteccia parietale sinistra danneggiano solo “il tempo oggettuale” ed il “tempo relazionale”
Lesioni nell’area visiva primaria oppure al confine tra l’area visiva primaria e la corteccia parietale, possono danneggiare in qualsiasi punto avvengono, tanto lo “spazio oggettuale” e lo “spazio relazionale” in costruzione quanto il “tempo oggettuale” e il “tempo relazionale” in costruzione.
Ciò è dovuto al fatto che, a questi livelli di elaborazione dei segnali provenienti dalla retina, il sistema è ancora organizzato retinotopicamente.
Affermiamo che, a questo livello, i due spazi e i due tempi agiscono a livello retinotopico e sono presenti in tutto il campo visivo, ripartito nei due emicampi destro e sinistro.
Avremo così che, per la visione, una lesione destra a livello occipito/parietale, dove lo spazio ed il tempo sono strutturati retinotopicamente può causare danni tanto alla parte destra dello “spazio” (oggettuale e posizionale) e del “tempo” (oggettuale e “relazionale), analogamente una lesione sinistra a livello occipito/parietale dove lo spazio ed il tempo sono strutturati retinotopicamente, può causare danni tanto alla parte sinistra dello “spazio” (oggettuale e posizionale) e del “tempo” (oggettuale e “relazionale)
Disorientamento visivo
Una lesione che interessa la costruzione degli sfondi mobile e fisso a livello retinotopico (i due spazi oggettuale e posizionale e i due tempi oggettuale e relazionale) nella regione occipito/parietale, ha come conseguenza la patologia nota con il termine di “Disorientamento visivo” Essa fu spiegata da Holmes (1918) .
“Egli descrisse pazienti che non riuscivano a localizzare la posizione o la distanza degli oggetti nello spazio solo per mezzo della vista. Non erano in grado di raggiungere gli oggetti, né di valutare le relative dimensioni di oggetti nel loro campo visivo. Un paziente (Private M.) ad esempio commetteva errori quando gli veniva chiesto quale di due oggetti fosse più vicino a lui e aveva parecchi problemi anche quando i due oggetti erano separati di 10 o 15 cm, a una distanza di mezzo metro da lui. Egli commentava: “ Quando guardo un oggetto mi sembra di andare più in là. Quando cerco di vedere qual è il più vicino sembra che cambino posizione in ogni momento: quello che guardo direttamente sembra scappare via”... I pazienti descritti da Holmes avevano anche difficoltà anche nel fissare i singoli oggetti, i loro occhi non convergevano su un bersaglio ed erano incapaci di seguire visivamente il movimento degli oggetti…” (
Questi pazienti non riescono ad eseguire le più semplici azioni quotidiane quali, per esempio, pranzare. Non riescono a localizzare correttamente il cibo nel piatto. La loro condizione spesso è peggiore di quella dei ciechi.
L’atassia ottica
“…Col termine di atassia ottica si intende un deficit consistente in una spiccata imprecisione di movimenti dell’arto superiore verso un obiettivo posto nel campo visivo, in assenza di un deficit sensitivo o motorio capace di spiegarne il sintomo.
Il disturbo può estendersi a tutto il campo visivo o può essere localizzato ad un solo emicampo (il destro o il sinistro). Può interessare entrambi gli arti superiori o un solo arto. Seguendo la classificazione di Rondot e coll (1977) , si parla di atassia visuo-motoria unilaterale quando il disturbo è localizzato ad un solo emicampo visivo. Nell’atassia visuo-motoria bilaterale il deficit interessa invece l’intero campo visivo. In entrambi i casi il disturbo può colpire tutte due le mani o una sola mano. Nel caso riguardi la mano omolaterale dell’emicampo esaminato si parla di atassia diretta, quando, invece è interessata la mano controlaterale all’emicampo esaminato si parla di atassia crociata …Mentre l’atassia ottica nella scimmia è limitata all’arto controlaterale della lesione, nell’uomo il fatto che il deficit sia confinato a un arto o a un emicampo spaziale sembra dipendere dal lato emisferico della lesione: e così, se la lesione è a destra si osserva un’atassia di entrambi gli arti superiori nel raggiungimento di obiettivi posti nell’emicampo sinistro, mentre se la lesione è a sinistra, si osserva un deficit della mano destra in entrambi gli emicampi e un deficit della mano sinistra limitato all’emicampo controlaterale”.
I pazienti affetti da questa patologia sono in grado di fissare stimoli visivi (non sono affetti da disorientamento visivo) e sanno indicare in modo appropriato stimoli tattili ed uditivi. Hanno difficoltà nel raggiungere oggetti sotto la guida visiva. Il difetto non è puramente visivo né esclusivamente motorio, piuttosto è un difetto di coordinazione tra i due campi.
L’atassia ottica è una patologia retinotopica e somatotopica; essa riguarda gli emicampi visivi destro e sinistro organizzati retinotopicamente, nonché gli arti destro e sinistro organizzati somatotopicamente.
Per comprendere questa patologia, si deve considerare il fatto che prendere un oggetto con la mano comporta due consapevolezze.
La prima concerne il mondo fisico dell’oggetto osservato con il suo spazio oggettuale e relazionale.
La seconda consapevolezza riguarda il mondo psichico dell’arto con le sue contrazioni muscolari ordinate nel tempo.
Rispetto al movimento ad occhi chiusi, non finalizzato al raggiungimento di un oggetto nello spazio, cambia lo spazio fisico e l’oggetto fisico.
Per prendere un oggetto, per esempio una mela, infatti, le contrazioni muscolari devono adattarsi allo spazio “oggettuale” e allo spazio “relazionale” della mela.
Lo spazio “oggettuale” della mela corrisponde alla componente distale del movimento (fase di manipolazione) e cioè la tendenza della mano ad assumere in anticipo una conformazione adatta all’oggetto da afferrare. Lo spazio “relazionale” della mela, corrisponde alla componente prossimale del movimento (fase di trasporto), che si manifesta nella precisione della direzione del movimento.
A mio avviso, avviene che lo spazio visivo a livello occipito/parietale ripartito retinotopicamente nei due emicampi destro e sinistro occupa il posto dello spazio propriocettivo. Ciò avviene nelle aree del lobulo parietale superiore e del solco interparietale (interessate dall’atassia ottica) (Perenin e Vighetto 1988) (Pierrot-Desailligny e coll. 1986)
In dettaglio, si può supporre che nell’emisfero destro proietta lo sfondo immobile dell’emicampo visivo sinistro, che si integra con la propriocezione dell’"arto superiore destro” . Separatamente, nello stesso emisfero, proietta lo spazio dell’emicampo visivo sinistro”, integrato con la propriocezione dell’arto superiore sinistro”.
Diversamente accade nell’emisfero sinistro nel cui lobulo parietale superiore e solco interparietale proiettano “spazi” di entrambi gli emicampi”, che si integrano con la propriocezione dell’arto superiore destro” e separatamente, lo “spazio” dell’“emicampo visivo destro”, integrato con la “ propriocezione dell’arto superiore sinistro”.
Ecco schematicamente le associazioni nel lobulo parietale superiore e nel solco interparietale dell’emisfero destro degli esseri umani.

Il seguente modello schematizza le associazioni presenti nel lobulo parietale inferiore e nel solco interparietale dell’emisfero sinistro umano

Agnosia per la forma
Affinché un oggetto sia separato dallo sfondo è necessario il buon funzionamento del sistema occipito/parietale, per la ovvia ragione che la spazio oggettuale e lo spazio posizionale sono importantissimi in questo compito. Analogamente se bisogna discernere le figure che in un disegno appaiono sovrapposte, oppure differenziare figure irreali ma possibili da figure irreali ma impossibili, è fondamentale l’attività della corteccia occipito/parietale.
L’insieme di queste patologie rientrano, secondo gli studiosi nella categoria di “agnosia per la forma”.
Come vedremo in seguito trattando delle agnosie legate alle patologie occipito/temporali, la “agnosia per la forma” ha ulteriori modalità di manifestazioni.
LE PATOLOGIE DELLA CORTECCIA PARIETALE
Per quanto riguarda la visione e il movimento volontario nella corteccia parietale destra e precisamente nel lobulo parietale inferiore gli sfondi fissi dei due emicampi destro e sinistro si integrano in un unico sfondo che avvolge l’intero oggetto.
Nella corteccia parietale sinistra e precisamente nel lobulo parietale inferiore, gli sfondi mobili dei due emicampi, destro e sinistro, si integrano in un unico sfondo che avvolge l’intero oggetto.
Il primo, con le figure in esso incollate è il sistema di riferimento spaziale; il secondo con le figure in esso incollate è il sistema di riferimento temporale
Un danno al primo procura l’eminegligenza spaziale unilaterale; un danno al secondo determina l’aprassia ideomotoria.
La negligenza spaziale unilaterale
“Si tratta di una sindrome caratterizzata da una alterata rappresentazione del contenuto del lato sinistro (più frequentemente) o lato destro dello spazio. Il paziente affetto da tale patologia si comporta come se non fosse più in grado di percepire e concepire l’esistenza del lato sinistro dello spazio egocentrico, corporeo ed extracorporeo.
Nella lettura di parole o titoli di giornali la NSU si manifesta come amputazione del segmento sinistro dello scritto e, spesso, nel caso di lettura di parole, come completamento patologico: il paziente sostituisce al segmento omesso un frammento inventato, che dà tuttavia luogo, di regola, a una parola realmente esistente nel suo lessico.
Invitato a indicare il punto centrale di una linea orizzontale il paziente lo pone più o meno spostato verso destra
Nel disegno di una figura elementare come una margherita o il quadrante di un orologio vengono omessi petali ed ore nel lato sinistro.
Manifestazioni di NSU possono essere presenti indipendentemente dal controllo visivo. Il paziente, ad occhi chiusi non riesce a toccare con la destra la propria mano sinistra. Per quanto riguarda la sfera extracorporea, nella ricerca cieca di oggetti sparsi sulla superficie di un tavolo, il paziente tralascia quelli posti a sinistra. Nella modalità uditiva, l’ammalato disloca verso destra uno stimolo acustico dicotico (Altman e coll., 1978) (Bisiach e coll 1984) .
Fenomeni di NSU possono essere rilevati, indipendentemente da qualunque stimolazione sensoriale, nella sfera della pura rappresentazione mentale. Si chiede al paziente di formare l’immagine visiva di una determinata configurazione (una stanza del proprio appartamento), secondo una prospettiva ben precisa e rigida, e di descriverne i dettagli. Nel fare ciò il paziente può omettere particolari salienti localizzati a sinistra. Se immediatamente dopo gli si chiede di ridescrivere la stessa configurazione secondo una prospettiva esattamente opposta alla prima, è possibile rilevare come i particolari precedentemente omessi in quanto localizzati a sinistra, vengano ricordati, una volta mentalmente trasferiti a destra e viceversa.
Nei pazienti affetti da NSU si possono manifestare dissociazioni cospicue, difficilmente spiegabili. Un paziente può invariabilmente leggere solo le ultime due lettere di parole che gli vengono presentate, anche se il foglio su cui sono scritte è posto completamente alla sua destra, mentre non compie alcun errore in una prova di indicazione di bersagli su un foglio delle stesse dimensioni. Un altro paziente può comportarsi in modo completamente opposto…
E’ possibile che dissociazioni di comportamento compaiono anche in prove interessanti diverse modalità sensoriali.
E’ importante sottolineare il fatto che la NSU si può presentare tanto con lesioni nella corteccia parietale destra (più frequentemente), quanto con lesioni nella corteccia parietale sinistra.
Tuttavia i pazienti destri si differenziano dai pazienti sinistri poiché i primi hanno danni nello sfondo fisso su cui è stampata la figura pregnante, i secondi hanno un danno nello sfondo mobile su cui sono incollate le figure scartate.

Facciamo alcuni esempi. In test di cancellazioni di linee, elaborato da Albert (1973) , i pazienti sono chiamati a cancellare in un gruppo di linee sparse in un foglio, ogni linea orientata diversamente.
Il 37% di pazienti con lesioni emisferiche destre ed il 30% di casi con lesioni sinistre omettevano una o più linee. C’era però una differenza nella distribuzione delle omissioni: i casi con lesioni destre tralasciavano più stimoli a sinistra, mentre i casi con lesioni sinistre facevano più errori casuali, omettendo di cancellare non solo stimoli a destra e al centro del foglio ma anche dimenticando stimoli a sinistra.
La spiegazione è la seguente. La lesione destra distrugge una porzione dello spazio “oggettuale” delle linee. Esse sono, quindi tagliate. Una lesione sinistra comporta difficoltà nella ricerca di linee sparse nello spazio. Questa ricerca è infatti attivata dal sistema occipito/temporo/parietale sinistro, dove è prevalente l’elemento temporale del “prima” e del “dopo”. Il paziente sbaglia perché è alterato l’oggetto psichico con la sua durata.
Altri studiosi hanno esaminato sistematicamente il ruolo del test nell’evidenziare la presenza di negligenza in seguito a lesioni destre o sinistre. Gainotti, D’Erme, Monteleone e Silveri (1986) condussero uno studio in cui doveva essere esaminato un gruppo di figure di animali allo scopo di indicare la presenza o l’assenza di un bersaglio presentato visivamente. Fu trovata un’incidenza di negligenza simile alle lesioni destre e sinistre. In un compito più complesso da un punto di vista percettivo (ricerca di un bersaglio in figure sovrapposte), il gruppo con lesioni destre risultava selettivamente compromesso perché tralasciava gli stimoli che si trovavano sul lato sinistro della figura composta. Questi studiosi ritennero che i pazienti con lesioni destre (o negligenza sinistra) mostravano il disturbo in presenza di quegli stimoli che potevano essere “visti con una singola fissazione”. Le lesioni sinistre davano origine a negligenza in compiti meno definiti che richiedevano ricerca attiva.
La nostra interpretazione è la seguente: Lo sfondo fisso o “spazio oggettuale” è fondamentale per la costruzione (il disegno) della figura.. Un danno allo sfondo mobile crea difficoltà nella costruzione della figura che viene tagliata
Al contrario un danno allo sfondo mobile della corteccia parietale sinistra impedisce la ricerca ordinata degli oggetti.
L’aprassia ideomotoria
“Si parla di aprassia quando un paziente non riesce ad eseguire un gesto richiestogli dall’esaminatore, benché non abbia difetti di moto, di senso, di coordinazione, che giustifichino il suo fallimento, o perché sono assenti in assoluto, o perché non interessano l’arto esaminato, che di solito è quello omolaterale alla lesione. Raramente l’aprassia appare nell’attività di tutti i giorni, benché la sua frequenza sia considerevole – dell’ordine del trenta per cento dei cerebrolesi sinistri – quando venga esaminata con prove ad hoc. Il fatto è che questo sintomo è contraddistinto al massimo da quella dissociazione automatico-volontaria sulla quale aveva richiamato l’attenzione Jackson a proposito dell’afasia. Lo stesso gesto che è eseguito correttamente in risposta a sollecitazioni contestuali o ad esigenze interiori, fallisce quando viene richiesto dall’esaminatore, fuori da ogni motivazione interna o esterna e quindi come produzione artificiale e altamente intenzionale…
La frattura nell’organizzazione gestuale può avvenire per due ragioni: o perché il paziente non sa cosa deve fare, non riesce, cioè, a rappresentarsi mentalmente il gesto da compiere, o perché egli non è in grado di tradurre la sequenza motoria che ha in mente in un corretto programma innervatorio, non sa, cioè, come fare. A questi due tipi di disturbo corrispondono, rispettivamente, l’aprassia ideativa (AI) e l’aprassia ideomotoria (AIM)”
L’aprassia ideomotoria insorge frequentemente dopo lesioni del lobulo parietale inferiore sinistro .
Abbiamo scritto in precedenza che il movimento di una parte del nostro corpo, per esempio la mano, comporta la consapevolezza del mondo psichico dell’arto con le sue contrazioni muscolari ordinate nel tempo. Questa consapevolezza è dominante rispetto alla consapevolezza dell’oggetto fisico collocato nello spazio. Per tale motivo, quando ci muoviamo nello spazio che ci circonda la “percezione” degli oggetti e la loro localizzazione è subordinata alla consapevolezza della “durata” del movimento e delle sue relazioni temporali.
In fondo, muovere una mano in vari punti di una stanza è come effettuare una ricerca con gli occhi di oggetti sparsi nella stessa stanza.
Lesioni al lobulo parietale sinistro, distruggendo lo sfondo mobile, danneggiano l’organizzazione temporale della ricerca oculare e della ricerca manuale.
L’autotopoagnosia
Descritta per la prima volta da Pick (1908) , l’autotopoagnosia. (perdita della conoscenza spaziale del proprio corpo) consiste in una difficoltà selettiva (riguarda solo le parti del corpo) ad indicare, sia su ordine verbale, che su imitazione, parti del corpo che vengono correttamente riconosciute una volta isolate da parte dell’esaminatore. Si tratta di una patologia abbastanza rara.
Per diagnosticare la seguente patologia si usano test che richiedono una mediazioneverbale e test non verbali.
Un esempio di test con mediazione verbale è l’invito volto al paziente di toccare su di sé una parte del corpo su comando verbale (Tocca l’orecchio sinistro!). Un esempio di test non verbale consiste nell’invito volto al paziente di toccare su se stesso la parte del corpo corrispondente ad un disegno mostrato dall’esaminatore.
L’autotopoagnosia si manifesta dopo lesioni tumorale della corteccia parietale sinistra.
Per spiegare questa patologia è stata proposta una ipotesi spaziale
“Se si considera il corpo come un insieme finito, l’isolamento di una parte specifica richiede la costruzione di un’immagine mentale della disposizione spaziale di come le varie parti siano poste in relazione tra di loro e nell’intero insieme: se tale capacità viene a mancare si osserverà una dissociazione tra riconoscimento e denominazione da una parte e indicazione e descrizione dei vari distretti corporei dall’altra. Tale dissociazione rappresenta la caratteristica principale dei pazienti autotopoagnosici
A differenza dei disturbi della conoscenza corporea da lesione emisferica sinistra, una lesione emisferica destra, può manifestarsi con grossolani disturbi della consapevolezza corporea, fino alla negazione esplicita dell’esistenza dell’emisoma (emisomatoagnosia).
Nelle attività di ogni giorno il paziente può comportarsi come se la metà sinistra del corpo non esistesse, tralasciando di rasare o truccare l’emifaccia sinistra o non infilando la stanghetta sinistra degli occhiali.”
Lesioni parietali all’emisfero sinistro impediscono, per i motivi già chiariti in precedenza, una ricerca attiva delle parti del corpo che non sono individuate.
Lesioni parietali dell’emisfero destro tagliano lo spazio “oggettuale” causando la cancellazione di una parte del proprio corpo.
“Kossilyn e collaboratori (1989) hanno dimostrato che i giudizi spaziali di tipo categoriale (sopra/ sotto, destra/ sinistra) vengono svolti più rapidamente quando gli stimoli sono presentati nell’emisfero sinistro, mentre le valutazioni delle distanze (giudizi in termini di coordinate visuo-spaziali) sono più rapidi per stimoli presentati all’emisfero destro.
E’ possibile che, quando consideriamo la destra o la sinistra di un oggetto o dello spazio nonché il sopra ed il sotto, effettuiamo una ricerca il cui elemento dominante è il tempo.
LA MEMORIA
Prima di illustrare la funzione e le patologie legate alla corteccia temporale (costruisce le due figure) è necessario chiarire, anche se per sommi capi, il funzionamento della memoria.
Di questo argomento ci occuperemo, nei dettagli, in un altro saggio.
Al fine della comprensione di quanto verrà di seguito espresso, possiamo affermare che la memoria è un ricostruire quanto è stato costruito in precedenza.
Quando, per esempio, vediamo un “oggetto”, per esempio il nostro cane, i recettori sensoriali della vista attivano i sistemi occipito/parietale ed occipito/temporale che costruiscono le due figure e i due sfondi. Quando ci “rappresentiamo mentalmente” l’immagine del nostro cane, alcune strutture subcorticali riattivano i centri corticali occipito/parietale ed occipito/temporale che ricostruiscono figure e sfondi.
La differenza tra “percezione” e “rappresentazione mentale” è determinata esclusivamente dai circuiti che attivano i sistemi del “dove” e del “che cosa” (dorsale e ventrale per la visione). Quando il circuito è costituito dai recettori sensoriali + il talamo si ha la “percezione”. Quando il circuito è costituito da ippocampo + corpi mammillari + talamo si ha la “rappresentazione mentale”.
L’ippocampo, i corpi mammillari ed il talamo hanno funzioni diverse.
Chiariamole con un esempio.
Supponiamo di guardare per la prima volta una persona, di nome Francesco. I recettori della retina inviano i loro segnali al “talamo”
Il talamo fa scaricare ordinatamente le colonne dei due sistemi ventrale e dorsale, in tal modo Francesco viene costruito all’interno dei sistemi di riferimento spazio/temporali.. I due sistemi, intanto, inviano impulsi all’ippocampo. Questa struttura subcorticale ha il compito di costruire un “codice di attivazione” . I corpi mammillari conservano questo codice di attivazione costruito dall’ippocampo sotto gli impulsi della corteccia.
Se il giorno dopo richiamo alla mente la “immagine” di Francesco, i corpi mammillari, utilizzando i loro codici di attivazione, inviano segnali al talamo che mette in funzione i due sistemi, “dorsale” per gli sfondi, “ventrale” per le figure. In tal modo la rappresentazione mentale è costruita. Rispetto alla percezione, la “rappresentazione mentale” è poco nitida, non essendo attivata dai recettori sensoriali della retina.
Il ricordo si differenzia dalla rappresentazione mentale in quanto esso ha la funzione di riconoscere immediatamente un oggetto e di integrare la percezione con l’esperienza vissuta.
Vediamo come funziona.
Supponiamo, dopo una settimana di incontrare Francesco. Ancora una volta i recettori della retina inviano i loro segnali al talamo che attiva le due vie del “dove” e del “che cosa”. I due sfondi e le due figure vengono in tal modo costruite.
Contemporaneamente si attiva il circuito subcorticale, corpi mammillari – talamo. Esso invia segnali alla corteccia affinché costruisca autonomamente le due figure. Queste due figure, immobile e mobile, contengono l’impronta degli sfondi di Francesco già percepiti, la settimana precedente. Lo sfondo, infatti, avvolge la figura, la quale ne è la parte mancante.
Sui due sfondi percepiti si inseriscono le figure conservate in memoria. In tal modo il cervello le riconosce.
Ovviamente è possibile il procedimento inverso. I circuiti subcorticali attivano gli sfondi su cui si inseriscono le figure costruite nella percezione.
Se ad essere danneggiata è la corteccia temporale è impedito il primo procedimento. Si ha in questo caso l’amnesia mnestica. Se ad essere danneggiata è la corteccia parietale viene impedito il secondo procedimento. Si ha in questo caso l’amnesia appercettiva.
Quanto abbiamo su detto sul ricordo è la spiegazione più semplice. In effetti è ovvio che il meccanismo debba essere più complesso. Basti pensare che, entrando nella nostra stanza da pranzo, con un breve sguardo ricostruiamo lo spazio circostante con gli oggetti presenti.
A mio avviso vi è una integrazione delle nuove esperienze con l’intera nostra conoscenza. Essa avviene sfruttando più figure e più sfondi costruiti dai due sistemi. Alcuni aiutano ad arricchire l’esperienza, altri costituiscono la semplice percezione. D’altronde si sa che tanto la via parvicellulare quanto la via magnocellulare sono formati da più vie parallele che proiettano nella corteccia.
Perché non supporre che ciascuna via costruisca una propria figura ed un proprio sfondo?
Questo spiegherebbe perché quasi tutte la patologie hanno numerose dissociazioni. La NSU, per esempio, può colpire selettivamente la rappresentazione mentale, essendo salvaguardata la percezione; oppure può accadere l’inverso.
LE PATOLOGIE DEL SISTEMA OCCIPITO/TEMPORALE
A livello occipito/temporale dove i costrutti sono organizzati retinotopicamente, le patologie interessano i primi stadi di costruzione delle due “figure” o “oggetti”, la prima legata al mondo fisico (emisfero destro); la seconda legata al mondo psichico (emisfero sinistro)
Agnosia per le forme
“Questa patologia identifica la incapacità di cogliere l’input sensoriale in una forma pregnante che si stacchi rispetto allo sfondo ed ad altre figure. Il paziente fallisce in compiti di accoppiamento di figure uguali, nel seguire con uno stilo i contorni di una figura, nel discriminare un quadrato da un rettangolo o un ellissi da un cerchio e non è in grado di copiare un disegno elementare”. (Con linguaggio realistico è descritta la funzione della corteccia occipito/temporale di costruire la figura.
L’agnosia per le forme fa parte della agnosia appercettiva. In genere questa agnosia concerne i primi stadi della percezione, quando la figura viene costruita a livello retinotopico
LE PATOLOGIE DELLA CORTECCIA TEMPORALE
l’agnosia associativa.
Questo tipo di agnosia concerne stati di elaborazione più avanzata del processo di costruzione delle figure.
Il termine associativa è, a mio avviso improprio. Come vedremo in seguito, le varie associazioni sono realizzate nella corteccia frontale per cui non vi è alcun legame tra una figura ed un’altra nella corteccia temporale. Io parlerei di integrazione dell’esperienza. Se, per esempio, entriamo in una stanza mai vista prima, questa immagine, attiverà, a livello subcorticale e, perché no, anche a livello percettivo, (le figure come abbiamo supposto sono più d’una) rappresentazioni di una conoscenza generale delle stanze. Sappiamo che hanno una forma di parallelepipedo, che alcune sono provviste di porte e/o finestre. Già la stessa luminosità ci fa comprendere, prima ancora della sua visione diretta, la presenza di un’apertura (porta o finestra) da cui filtra la luce. Fatto analogo avviene per quanto riguarda i vari oggetti, per esempio i televisori. I vari costrutti si sovrappongono costituendo una immagine generale che si attiva quando viene visto un nuovo televisore con il suo sfondo.
CANE
Consideriamo la seguente parola:
I due sistemi occipito/parietale ed occipito/temporale, possono, separando figure e sfondi costruire i seguenti oggetti

La parola “cane”, può costituire una singola figura, oppure l’insieme di più figure. Queste considerazioni sono però della corteccia frontale.
LA CORTECCIA FRONTALE
Compito della corteccia frontale è quello di associare i costrutti delle aree primarie temporale e parietale In tal modo costruisce i significati e quindi, per quanto riguarda l’uomo i pensieri
In che modo la corteccia frontale attua questo processo?
Supponiamo di osservare due oggetti posti nello spazio per esempio “Giovanni” ed una “cesta di pane”
I sistemi dorsale e ventrale costruiscono due sfondi e due figure. Abbiamo così due “oggetti fisici” e due “oggetti psichici”.
Consideriamo lo “oggetto fisico”
Il cervello costruisce la prima figura con lo sfondo; prima di costruire la seconda figura effettua una pausa attenzionale. Se così non facesse, infatti, non vedremmo figure separate ma un groviglio di immagini mischiate l’una all’altra. Dopo questa pausa costruisce il secondo oggetto. Possiamo rappresentare così questo processo:

Durante la pausa, lo sfondo viene attivato, altrimenti per una frazione di secondo il mondo fisico scomparirebbe nel nulla. Diciamo che la costruzione dello sfondo accompagna la costruzione delle due figure con la pausa interposta. Indichiamo lo sfondo fisico della corteccia parietale destra tramite una linea continua sopra le due immagini . La rappresentazione completa sarà:

Compito della corteccia frontale in questo costrutto è quello di associare le figure della corteccia occipito/temporale con lo sfondo della corteccia occipito/parietale.
La corteccia frontale, inoltre, tiene insieme tramite la memoria di lavoro i due oggetti con lo sfondo. Indichiamo con una doppia freccia la memoria di lavoro, avremo:

Il costrutto sopra raffigurato è il prototipo dei “verbi”. Si tratta di un passaggio da un oggetto ad un altro all’interno di uno sfondo che tiene insieme i due oggetti, il tutto unificato dalla memoria di lavoro.
Supponiamo che la corteccia frontale costruisca la prima figura con lo sfondo e dopo la pausa attenzionale tolga lo sfondo alla seconda figura. In questa circostanza la figura viene cancellata, se ne avrebbe una rappresentazione inconsapevole. Possiamo raffigurare questa circostanza così:

Il passaggio verbale è, in questo caso eliminato. Questa è la struttura di tutti i sostantivi.
Se ad essere eliminato è lo sfondo della prima figura avremo:

Abbiamo in questo caso la struttura di tutti gli aggettivi
Combinando in vari modi figure e sfondi di oggetti fisici e psichici la corteccia frontale costruisce i molteplici significati nonché i pensieri.
Per esempio il pensiero “Giovanni curioso” potrebbe corrispondere alla struttura:

Corrispondenza tra le formule di Vaccarino e gli schemi illustrati
In linea generale gli schemi soprastanti rispecchiano le formule di Vaccarino. Vi sono tutti gli elementi essenziali e cioè i momenti attenzionali, applicati e puri, la memoria strutturante e la memoria di lavoro.
Possiamo supporre che il pensiero, associandosi al linguaggio e quindi ai suoni, possa svincolarsi dalla “realtà esterna” ed essere costituito esclusivamente da momenti attenzionali puri (nel nostro esempio il cerchietto).
Si può pensare che, per fare ciò, la corteccia frontale dovrebbe semplicemente attivare le figure senza lo sfondo.
Ciò però non è possibile. Infatti, venendo a mancare lo sfondo che costituisce la memoria strutturante, non avremmo le formule.
Dobbiamo allora supporre una precisa area della corteccia cerebrale dove l’attivazione di figura e sfondo sia fittizia, non applicata cioè a percezioni sensoriali. In questo caso avremmo un’area per le categorie e altre aree per i costrutti sensoriali.
Un’area associata al movimento potrebbe svolgere questa importantissima funzione. Sono comunque necessarie ricerche ad hoc.
E’ comunque possibile ottenere le formule di Vaccarino, utilizzando gli schemi fin qui proposti.
Sono però necessarie alcune considerazioni.
La corteccia frontale associando sfondo e figura ha le seguenti possibilità:

inserire la figura nello sfondo:

2) prendere solo la figura, senza lo sfondo:

prendere lo sfondo senza figura
Supponiamo che la corteccia frontale agisca su due sistemi occipito/parietale ed occipito/temporale che costruiscono figura e sfondo. A tal proposito c’è da sottolineare il fatto che a livello talamico, per ciascun emisfero vi sono due vie magnocellulari nonché quattro vie parvicellulari, due delle quali funzionali per la percezione dei colori.
Per differenziare i costrutti dei due sistemi utilizziamo due linee, una tratteggiata ed una continua.
Adesso rappresentiamo sulla sinistra le formule di Vaccarino e sulla destra gli schemi corrispondenti alle figure/sfondi dei due sistemi

(Paolo Nichelli (1996): Disturbi spaziali e visuo-immaginativi. Manuale di neuropsicologia 515 - 517
Edoardo Bisiach: Negligenza spaziale unilaterale e altri disordini unilaterali di rappresentazione Manuale di neuropsicologia 639-661 (1992).
Zeki S.M. (1974) Functional specialization of a visual cortex and its relation to visual agnosias in the posterior bank of the superior temporal sulcus of the thesus monkeys. Journal of Phisiology,236: 549 - 573
Mishkin M., Ungerleider L. G., Macko K. A. (1983) Object vision and spatial vision: Two cortical pathways. Trends in Neuroscience, 6, 414 - 417
Livingstone M., Hubel D. (1988) Segregation of form, colour, movements and depth: Anatomy, physiology and perception: Science, 240: 740 - 749
Perenni M. T., Vighetto A. (1988) Optic Ataxia: A specific disruption in visuo-motor mechanisms, 1. Different aspects of the deficits in reaching for obiects. Brain, 111: 643-674
Pierrot-Deseilligny C., Gray F., Brunet P., (1986) Infarcts of both parietal inferior lobules with impairment of visually guided eye movements, peripheral visual inattention and optic ataxia. Brain 109: 81-97
Altman J.A., Balonov L.J., Deglin V.L., (1978) effects of unilateral disorder of the brain hemisphere function in man on directional hearing. Neuropsycologia, 90: 21-26
Bisiach E., Cornacchia L., Sterzi R., Vallar G. (1984) Disorders of perceived auditory lateralization after lesions of the right hemisphere. Brain 107: 37-52
Gainotti G., D’Erme P., Monteleone D., Silveri M.C. (1986) Mechanisms of unilateral spatial neglect in relation to laterality of cerebral lesions. Brain, 109: 599-612
De Renzi E., Faglioni P., Lodesani M., Vecchi M., (1983) Impairment of left brain-damaged patients – on imitation of single movements and motor sequences – frontal and parietal injured patients compared. Cortex, 19: 333-343
Pick A. Uber Storungen der Orienterung am einigen Korper. In Arbeiten aus der deutschen psychiatrischen Universitaetklinik in Prag, Berlin: Karger
Kosslyn S.M., Koenig O., Barret A., Backer Cave C., Tang G., Gabrieli J.D.E. (1989) Evidence for two types of spatial representations: Hemispheric specialization for categorical and coordinate relations. Journal of Experimental Psychology: Human Perception and Performance, 15: 723-735.
Autore: Leonardi Salvatore
Fonte: http://www.methodologia.it/wp/Leonardi_136.doc
Link sito web: http://www.methodologia.it
IL CERVELLO QUESTO SCONOSCIUTO
Il cervello è un organo la cui complessità non ha confronto in tutte le cose note nell’universo, ma pur sempre composto di cellule come tutti i tessuti.
Si tratta di cellule altamente specializzate che obbediscono alle stesse leggi che governano le
altre cellule.
I loro segnali elettrici e chimici possono essere rilevati, registrati ed interpretati così come possono essere identificati i loro componenti chimici.
Si può costruire una “ carta geografica “ delle connessioni che costituiscono il reticolo fibroso del cervello. Il cervello, cioè, può essere studiato proprio come il rene o il fegato.
Il cervello è da molto tempo oggetto di ricerca scientifica e negli ultimi 15 anni la neurobiologia è diventata una delle branche più attive della scienza.
Nonostante questo, la ricerca sul cervello è solo agli inizi poiché la sua complessità é un dato di fatto.
Il cervello è una parte del sistema nervoso E in questo sistema ci sono due tipi di cellule: le cellule neurogliali e le cellule nervose o neuroni.
Le cellule neurogliali occupano tutto lo spazio non occupato dai neuroni. La loro funzione non è ancora del tutto nota ma sicuramente servono da “sostegno” alla delicata rete di neuroni.
I neuroni sono i “mattoni” di cui è fatto il cervello. Hanno caratteristiche uniche, grazie alle quali il cervello funziona in modo assai diverso, per esempio, dal fegato.
Il numero di neuroni che formano i circa 1400 grammi del cervello umano è dell’ordine di 10¹¹ ( = 100 miliardi ) una cifra all’incirca pari al numero delle stelle che si trovano nella nostra galassia !!
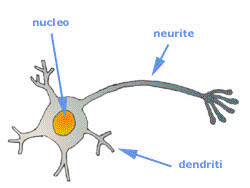
Un neurone è costituito da un “corpo cellulare” con un diametro compreso fra i 5 e 100 micrometri (= millesimi di millimetro).
Da questo fuoriescono una fibra principale chiamata “assone” e un gran numero di ramificazioni fibrose, chiamate “dendriti”
I circa mille miliardi di neuroni del sistema nervoso (di cui dieci miliardi nella corteccia) formano, pertanto, circa un milione di miliardi di contatti.
Il funzionamento del cervello dipende dal flusso di informazioni che passano in complessi circuiti formati da “reti ” di neuroni.
In questa complessa rete, lo scambio di informazioni tra neuroni avviene con modalità elettriche e con modalità chimiche.
Certe condizioni chimiche nel corpo del neurone generano una differenza di potenziale elettrico, sul segmento iniziale dell'assone. Questa tensione, una volta nata, si propaga sull'assone e, arrivata alle terminazioni, provoca l'invio di sostanze chimiche ai neuroni successivi. Tra neurone e neurone non c'è un collegamento diretto ma resta un piccolo spazio (sinapsi), dove transitano delle sostanze chimiche che realizzano il collegamento tra neuroni (neurotrasmettitori)
L’arrivo di queste sostanze è una informazione per il neurone che le riceve, per cui si parla di segnale in circolazione. Il segnale è elettrico dentro al neurone e chimico tra neurone e neurone (con l’eccezione di alcune sinapsi, dette sinapsi elettriche, dove il segnale elettrico transita direttamente).
I neurotrasmettitori possono essere sostanze eccitatorie o inibitorie. Per eccitare un neurone, cioè per far nascere una tensione su di esso, non è sufficiente l'arrivo di una sostanza eccitatoria da un altro neurone ma occorre l’azione di molti neuroni ad esso collegati per indurne l'eccitazione. Se poi gli arrivano anche neurotrasmettitori inibitori essi annullano una parte di quelli eccitatori. Un neurone si eccita solo se le eccitazioni ricevute superano le inibizioni di una quantità prefissata (funzionamento a soglia).
Se eccitato opportunatamente, dunque, il neurone modifica le sue caratteristiche fisiologiche ed è in grado di produrre segnali elettrici che si propagano nei suoi prolungamenti dendritici e assonici.
Le sinapsi quando vengono investite da questa carica elettrica provvedono alla conversione di tale segnale in segnale chimico.
Nella parte terminale della cellula presinaptica sono presenti delle vescicole che contengono dei mediatori chimici. Eccitate dall’impulso elettrico queste vescicole liberano il loro contenuto nello spazio sinaptico e tali sostanze si combinano con recettori chimici presenti nella cellula postsinaptica.
L’informazione viene, in questo modo, trasferita da una cellula ad un’altra grazie a questi particolari “punti di contatto” .
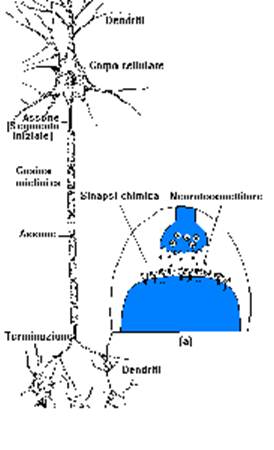
Un neurone può possedere da 1.000 a 10.000 sinapsi e può ricevere informazioni da migliaia di altri neuroni !
Un pezzettino di cervello grosso come la capocchia di un fiammifero contiene circa 1 miliardo di connessioni, senza considerare gli svariati modi in cui queste si potrebbero combinare!
Nell’organizzazione neurologica del sistema nervoso centrale umano i raggruppamenti di neuroni formano organi funzionali specifici, che si sono evoluti nel corso dell’evoluzione della specie.
Il midollo spinale è la struttura filogeneticamente più antica del nostro sistema nervoso.
Con lo sviluppo di altri organi nervosi sovrapposti al midollo spinale, questo ha acquistato la funzione di trasmettere le informazioni sensoriali ( tattili, dolorose, cinestesiche) al cervello e di indirizzare i comandi motori del cervello verso la periferia.
Una struttura antica come il midollo spinale è il cervelletto che controlla le attività motorie e cinestesiche che sono alla base dell’equilibrio, dell’orientamento spaziale, dei riflessi e della nostra postura eretta.
Midollo spinale e cervelletto formano il sistema spinomidollare.
Sovrapposto a questo si trova il paleoencefalo nel quale sono presenti organi come il talamo, l’ipotalamo, il mesencefalo ed altri...
Questi presiedono le funzioni emozionali ed emotive del comportamento. Nel talamo, per esempio, i segnali in arrivo si collegano a sentimenti di gioia, dolore, paura, piacere.
Così le informazioni in ingresso vengono confrontate con le esperienze precedenti e valutate in termini affettivi e ritrasmesse in altre aree cerebrali.
L’ipotalamo regola varie funzioni vitali come la fame, la sete, il sonno e la veglia.
Nel mesencefalo si verificano le prime decodificazioni sensoriali di vista ed udito.
La corteccia rappresenta l’ultimo stadio dell’evoluzione, chiamata per ciò anche neoencefalo.
I comportamenti regolati dal paleoencefalo non sono sempre coscienti, mentre le attività controllate dalla corteccia sono associate ad esperienze conoscitive e a processi volontari coscienti.
La corteccia è costituita da tre aree:
1) Le aree motorie : da qui partono informazioni nervose che producono movimenti
2) L’area sensoriale : qui arrivano le informazioni rilevate da tutti gli organi di senso
3) Le aree associative sono responsabili :
di associare tra di loro le informazioni dando loro un senso compiuto,
di discriminare ed interpretare i segnali
dell’attenzione, concentrazione, ....
Un po’ dappertutto nel sistema nervoso si trova un altro tipo di cellule, le cellule Schwann.
Tutti gli assoni sono ricoperti da cellule Schwann dando luogo a strati di isolante noti con il nome di mielina.
Questa “guaina” di mielina serve per conservare l’energia del neurone e per condurre gli impulsi elettrici più velocemente.
Il cervello inoltre è collegato al mondo esterno mediante neuroni specializzati, chiamati trasduttori sensoriali. Questi formano gli organi di senso e forniscono al cervello i segnali di ingresso, mentre i segnali di uscita passano attraverso i neuroni collegati ai muscoli e alle ghiandole.
Una porzione di tessuto cerebrale è una rete intricata che reagisce a segnali elettrici e chimici e mediante specifici collegamenti e connessioni influenza l’azione di altri organi del nostro corpo come il cuore, i reni, i polmoni, i muscoli.........
Il cervello è quindi un controllore generale che può, con i suoi schemi, modificare la respirazione, il battito cardiaco, la digestione, il movimento, ecc.
Come centro di decisione il cervello deve quindi conoscere in ogni momento quello che succede :
1) In casa sua: l’informazione viene dall’interno. Se tutti gli apparati interni funzionano bene essi non si manifestano alla coscienza anche in stato di veglia.
2) Fuori casa: qui le linee esterne trasmettono al cervello il flusso di informazioni provenienti da cinque ricettori specializzati. Il loro messaggio sensoriale ha 2 funzioni :
- Avvertire il cervello di ogni modifica dell’ambiente esterno e generare un adattamento locale o generale alla nuova situazione.
- Arricchire il contenuto cerebrale rispetto alle informazioni da memorizzare, ma anche stimolare l’organizzazione delle strutture e delle connessioni nervose.
Le informazioni che arrivano dall’esterno o dall’interno del corpo confluiscono tutte alla corteccia cerebrale.
In breve:
- c’è un segnale d’ingresso: l’unico mezzo con il quale l’uomo può conoscere il mondo esterno;
- c’è un segnale in uscita : l’unico modo con cui l’uomo risponde al mondo esterno e influisce su di esso.
Tra il segnale in entrata e il segnale in uscita c’è tutto il resto, percezione, emozioni, pensiero e tutto ciò che ci rende ESSERI UMANI .
Fonte: http://scg.itcilo.org/AreaPubblica/OrientamentoMobilita/Cognizione/cervello/attach/Cervello.doc
Autore del testo: non indicato nel documento di origine
Cervello umano
IL CERVELLO COME MACCHINA QUANTISTICA
Nel 1994 il più eminente fisico teorico dell’Università di Oxford, Sir Roger Penrose, pubblica Shadows of the Mind, discusso libro in cui afferma che il cervello elabora quantisticamente l’informazione grazie alla presenza di microscopiche molecole di tubulina all’interno dei neuroni.
Il cervello viene visto come una macchina quantistica di primo livello.
Il suo modello richiede che 20.000 neuroni si mantengano per 1/40 di secondo in sovrapposizione quantistica a temperatura ambiente.
Il suo lavoro ha molti detrattori. Sembra improbabile che in queste condizioni le peturbazioni dell’ambiente non danneggino la sovrapposizione.
Eppure se la vita ha imparato a sfruttare il livello quantistico, la temperatura ambiente deve potenziarne le prorietà e non annullarle.
L’idea fondamentale di Penrose deriva dall’osservazione che anche gli organismi unicellulari hanno comportamento finalizzato, reagiscono alla luce, evitano ostacoli, hanno una forma di memoria, senza avere neuroni.
L’agente primario del comportamento “intelligente” del paramecio sono le sue ciglia:
filamenti che sono sia antenne che strumenti per il movimento
elaborano informazioni dall’ambiente
generano i movimenti sincroni necessari per attuare le decisioni conseguenti
Le ciglia generano onde oscillatorie sincronizzate, pervenendo ad un’autoorganizzazione globale sulla base di interazioni locali.
Le ciglia del paramecio sono microtubuli.
I fusi mitotici che nelle cellule attuano “spontaneamente” la divisione e la ricomposizione in cellule figlie sono microtubuli.
La coda dello spermatozoo, che ondeggia muovendosi in direzione dell’ovulo, è una struttura a microtubuli.
I neuroni contengono molti microtubuli.
Probabilmente l’elemento fondamentale dell’intelligenza cellulare è il microtubulo.
Stuart Hameroff, medico ricercatore dell’Università dell’Arizona, negli anni 80 ipotizza che i microtubuli siano capaci di calcolo. Fino ad allora erano sempre stati considerati solo componenti del citoscheletro.
Nel tempo si è scoperto che le funzioni dei microtubuli sono:
- Il trasposrto intracellulare di organelli
- Il movimento ciliare, ondulatorio e sincronizzato
- L’organizzazione della divisione cromosomica (movimento oscillatorio sincronizzato)
- Scambio di segnali da un elemento all’altro
- Comunicazione fra nucleo e interno della cellula
- Crescita di collegamenti fra neuroni.
L’energia dei microtubuli è di tipo elettromeccanico, e la rete di microtubuli si comporta come un calcolatore elettromeccanico.
Hameroff ipotizzò nel 1982 che i microtubuli si comportassero come automi cellulari.
In realtà la struttura del microtubulo assomiglia esattamente ad un automa cellulare.
Ogni tubulo è costruito sulla base di migliaia di dimeri di tubulina sistemati in una griglia bidimensionale arrotolata su se stessa.
La griglia forma un vicinato esagonale con sei vicini.
Le cariche elettrostatiche provocano l’allineamento dei dimeri.
Il passaggio di un elettrone cambia la conformazione del dimero.
In un automa cellulare è necessario un “meccanismo di temporizzazione” che faccia scattare l’aggiornamento simultaneo delle celle: è necessario trasmettere un segnale regolare, una vibrazione periodica.
Oggi sappiamo che le reti di microtubuli trasmettono davvero forze fluttuanti, mantenendo quasi istantaneamente in tutta la rete un equilibrio dinamico.
Il meccanismo di temporizzazione può essere dato dalle onde ritmiche.
Tuttavia il modello di Hameroff era sbagliato: le regole che governano le interazioni fra tubuline non sono quelle da lui predette.
Inoltre il suo modello per funzionare necessita dello zero assoluto.
Le reti di tubuline assomigliano ad automi cellulari ma non lo sono.
Tuttavia abbiamo visto che funzionano davvero come computer paralleli. Il modo in cui calcolano è ancora da stabilire.
Il modello di Jack Tuszynski (Università di Alberta) è più convincente.
Parte dal principio che le reti di microtubuli devono funzionare a temperatura ambiente e dimostra che sono trasmettitori elettromeccanici di segnali che computano in base a un modello simile ai vetri di spin.
Il microtubulo funziona quindi come un cavo intelligente autoadattativo.
In questo caso la rete neurale del cervello è abbinata alla rete dei microtubuli e viceversa.
Un gruppo di scienziati della Wayne State University sta simulando un computer molecolare basato su microtubuli, che apprende come una rete di Hopfield e si basa sulla natura oscillatoria dei suoi elementi.
Si può a questo punto ipotizzare la natura quantistica di questi processi: infatti la tubulina, che possiede due possibili stati, a e b, passa dall’uno all’altro a causa del passaggio per effetto tunnel dal dimero a a quello b.
Fonte: http://www.dti.unimi.it/pizzi/CORSO%20QNN/5%20-%20IL%20CERVELLO%20COME%20MACCHINA%20QUANTISTICA.doc
Autore del testo: non indicato nel documento di origine
Cervello umano
Visita la nostra pagina principale
Cervello umano
Termini d' uso e privacy