Neurobiologia cellulare
Neurobiologia cellulare
Questo sito utilizza cookie, anche di terze parti. Se vuoi saperne di più leggi la nostra Cookie Policy. Scorrendo questa pagina o cliccando qualunque suo elemento acconsenti all’uso dei cookie.I testi seguenti sono di proprietà dei rispettivi autori che ringraziamo per l'opportunità che ci danno di far conoscere gratuitamente a studenti , docenti e agli utenti del web i loro testi per sole finalità illustrative didattiche e scientifiche.
Neurobiologia cellulare
Appunti del programma di Elementi di Neurobiologia cellulare
Sistema nervoso centrale ---> cervello e midollo
Sistema nervoso periferico ---> nervi e cellule nervose
Il cervello umano ha circa 1000 miliardi di cellule
I neuroni sono le uniche cellule a contribuire alla "vita mentale" perché hanno un'attività elettrica (impulsi nervosi) che sono alla base di una forma chimica/elettrica di calcolo. Le reti neurali fanno calcoli (elaborano le informazioni).
Come è composto un neurone? Corpo cellulare (soma) al cui centro vi è il nucleo (con i filamenti di DNA). Membrana esterna composta da lipidi, che contiene al suo interno acqua (citosol) in cui ci sono composti chimici necessari al processo di sopravvivenza (ciclo di Krebs)---> in presenza di glucosio e ossigeno fornisce energia (ATP). I prodotti di scarto sono anidride carbonica e acqua.
Metabolismo cellulare ---> metabolismo ossidativo (Krebs)
Sintesi proteica ---> Le proteine sono essenziali alla cellula, gli enzimi facilitano le reazioni chimiche. Le proteine possono essere______
La sint. prot. avviene nel reticolo endoplasmatico ruvido (con ribosomi) con RNA ribosomiale (RNA = acido ribonucleico). Una proteina viene sintetizzata all'interno del reticolo endoplasmatico ---> il filamento di DNA (amminoacidi) scorre lungo il ribosoma e produce la proteina.
La cellula nervosa produce neurotrasmettitori ---> l'apparato del Golgi produce i granuli secretori che si muovono lungo l'assone e diventano neurotrasmettitori. (il tetano fa in modo che vengano secreti in continuazione i neurotrasmettitori indebolendo l'organismo).
Trasporto assonale anterogrado ---> Le vescicole si muovono verso la sinapsi
Trasporto assonale retrogrado ---> Le vescicole tornano all'apparato del golgi (è sempre veloce)
Anterogrado lento = 1-10 mm al giorno
Anterogrado rapido = 1000 mm al giorno (con l'aiuto dei microtuboli)
Microtuboli = Stanno lungo l'assone e le vescicole si attaccano a loro x viaggiare più velocemente.
Retrogrado ---> la Rabbia (il virus) viaggia dalle sinapsi periferiche al cervello, attraverso questo meccanismo di ritorno.
Esocitosi ---> la vescicola sinaptica si fonde col neurone (la membrana)
Endocitosi ---> la membrana della vescicola viene risucchiata e torna indietro VEDI DISEGNO!
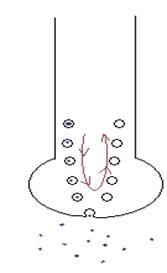 |
Output = il corpo elettrico viaggia verso l'esterno (dal soma all'assone alla sinapsi)
Input = attraverso i dendriti (propaggini multiple che raccolgono segnali in entrata)
Sinapsi asso-assoniche ---> che arriva sull'assone (non passa per il nucleo)
sin asso-dendritiche ---> che arriva dai dendriti
sin. somiche ---> che arriva nel soma
|

La mielina (Vedi dis.) avvolge l'assone e funge da isolante elettrico x velocizzare il passaggio dell'informazione nel sist. nervoso periferico.
Nel sist. nervoso centrale al posto della mielina c'è l'oligodendrocita, che fornisce mielina per svariati assoni. La sclerosi multipla compromette gli oligodendrociti che non producono più mielina e l'efficacia della trasmissione dei segnali del cervello si riduce e sparisce (avviene anche la paralisi).
Cellule glioli:
microglia ---> mangia virus e batteri (fagocita)
macroglia ---> sono molto più grandi (quanto i neuroni) e sono Oligodendrociti; cellule di Schwann; Astrociti
Astrociti ---> hanno forma di stella; svolgono funzione di sostegno (riempiono lo spazio fra neurone e neurone, mantenendo la forma degli stessi e impedendone lo spostamento); hanno funzione macrofagica; agiscono come barriera (filtrano i metaboliti che arrivano al cervello ed eliminano quelli tossici: sono un filtro fra sangue e cervello (barriera ematoencefalica); fanno da rivestimento endoneurale (barriera fisica) VEDI DIS.
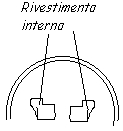 |
ventricoli laterali -> sono cisterne piene d'acqua nel cervello, il cui rivestimento interno è fatto da astrociti; assorbono gli ioni di potassio in eccesso; riassorbono i neurotrasmettitori in eccesso; hanno una funzione trofica (cercalo!)
Sostanza Bianca = il bianco è dato dalla mielina (assoni esterni)
Sostanza grigia = il grigio è dato dal soma (assoni interni)
Forza chimica ---> statisticamente le particelle si distribuiscono equamente
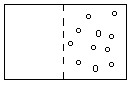 |
Forza elettrica ---> quando si perde un elettrone si ha ione positivo + (2 el. = ++, ecc..), se si guadagnano -
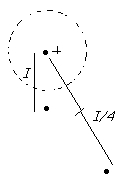 |
|
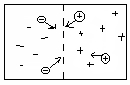
Un ione positivo ne respinge uno uguale | Le positive e negative si attraggono fino a mescolarsi.
con una forza i. |
Ione positivo = Catione
Ione negativo = Anione
Gradiente = zona in cui si devono distribuire le particelle
? Se ci sono molte particelle da una parte e pochissime dall'altra, per i primi passaggi gli spostamenti saranno tutto sommato uguali (variano di pochissimo).
Flusso ---> n° ioni / (msec x area)
forza x permeabilità membrana
Flusso chimico ---> forza chimica x permeabilità membrana
Flusso elettrico ---> forza elettrica x permeabilità membrana
Per flusso si intende sempre il flusso netto (50 e 50 è 0)
Adesso vedi slide n° BOH
Durante il potenziale di azione, c'è un'improvvisa permeabilità prima al sodio, poi si chiudono i canali per il sodio e dopo un po' si aprono quelli per il potassio.
Proteine nel neurone sono a forma di tubo, e hanno funzione selettiva: cioè far passare solo il sodio o il potassio (canali passivi)
Canali a voltaggio dipendente ---> ha due specie di membrane che si chiudono grazie a delle cariche positive che si spostano se c'è un potenziale negativo (spostandosi chiudono le membrane) o positivo (le aprono). Quindi quando il voltaggio supera una certa soglia si aprono.
Il canale a voltaggio dipendente per il sodio è più complicato. Secondo una teoria, questo canale ha due meccanismi di chiusura: 1) quello spiegato sopra, con le due cariche che si muovono a seconda del potenziale; 2) ci sarebbe una porzione globulare che tenderebbe a chiudere il poro; quando le membrane si aprono per via del potenziale elettrico, fluisce il sodio (per un brevissimo tempo, frazioni di secondi), poi la porzione globulare chiude il poro (fase di inattivazione)
Fase di inattivazione ---> il canale è chiuso,ma dalla porzione globulare. Questa fase è
attivabile ---> cioè se si torna ad un voltaggio basso le falde non si possono richiudere, perché bloccate. Il comportamento della porzione globulare è totalmente indipendente da quello delle falde.
Dopo un po' di tempo, la porzione globulare si stacca e, se il potenziale è basso, le falde si richiudono. VEDI FIG.
La fase di inattivazione è caratterizzata dalla refrattarietà assoluta ---> per quanto si depolarizzi quella zona, non può succedere nulla, non si può produrre un picco di ingresso.
Refrattarietà relativa ---> quando, dopo che la porzione globulare si leva, le membrane si richiudono. C'è bisogno di più potenziale elettrico.
- Il segnale va indietro perché c'è la refrattarietà assoluta
- L'apertura/chiusura di un canale a voltaggio dipendente dipende dal voltaggio (fenomeno - elettrico)
- La chiusura di un canale per via della porzione globulare non è un fenomeno elettrico
- La richiusura dipende dalla P.G. E dal potenziale elettrico sotto la soglia
- Il potassio ha un solo meccanismo di chiusura
- Il sodio ha due meccanismi di chiusura
- Durante la fase di refrattarietà relativa i canali sono chiusi
- iperpolarizzazione ---> quando si aprono i canali
- 0 ---> assenza di polarizzazione
- ripolarizzazione ---> fase di discesa in cui si ritorna al valore iniziale
Grazie ad una piccola depolarizzazione, la membrana si è spostata, senza nessun dispendio di energia; questo si propaga da membrana a membrana con spesa energetica 0.
L'unica spesa metabolica è quella per mantenere il potenziale a -60.
Tutto questo in 2-3 millisecondi, per un unico punto di una membrana.
Propagazione passiva o elettrotonica
Resistenza assiale ---> facilità con cui uno ione può affluire lungo l'assone (tanto più è grande l'assone, tanto più veloce andrà lo ione)
Resistenza membranosa ---> più è largo l'assone, più è probabile che gli ioni si distribuiscano lungo tutte le uscite; più è stretto, più si ammasseranno nelle prime uscite.
Per far si che si vadano più lontano possibile, si hanno due possibilità: 1) allargare l'assone; 2) isolare elettricamente le prime uscite con la mielina, che chiude le uscite.
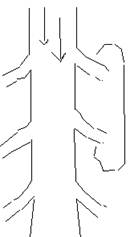 |
Quelli nella figura sono canali passivi. Questo sistema (mielinizzazione delle fibre) serve non solo per aumentare la distanza percorsa dagli ioni, ma anche a rendere più costante l'intensità.
â --> lamda = costante di spazio (+ â = più spazio nell'assone)
Nella propagazione passiva o elettrotonica, c'è la minimizzazione del tempo di uscita (gli ioni cercano di uscire più velocemente possibile); gli ioni sono tutti positivi e quindi si respingono. Sfrutta i canali passivi ---> non ha bisogno di energia.
Questa diffusione avviene ovunque, anche nei dendriti e nel soma, dove non ci sono i canali a voltaggio dipendente per il sodio.
Dendriti e soma ---> solo canali passivi
Assone e sinapsi ---> canali passivi + canali a voltaggio dipendente
Propagazione attiva:
- avviene solo se si supera la soglia, altrimenti non succede nulla;
- avviene solo con i canali a voltaggio dipendente per il sodio;
- non si esaurisce con la distanza (conserva la stessa intensità; non è decrementale)
Si basa sullo stesso meccanismo di quella passiva. Quando la soglia è superata, le membrane si aprono ed entra il sodio, con un'eccedenza di cariche positive all'interno e negative all'esterno. Nella parte vicina (a riposo) è il contrario con cariche negati all'interno e positive all'esterno. Gli opposti si attraggono e si forma un circuito elettrico, le cariche girano.
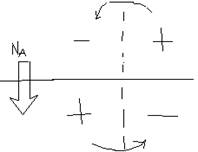 |
Le cariche positive tendono a spostarsi cercando di depolarizzare la zona a riposo (che è a -60 millivolt), questa aumenta, ma il potenziale d'azione si scatena anche nella zona a riposo. Le zone successive si attivano e questo “anello” si muove lungo l'assone, in direzione della sua fine, con la caratteristica di non perdere intensità lungo il tragitto (non come nei dendriti) e non consuma energia per via della natura elettrica degli ioni.
La propagazione attiva consiste nella diffusione della corrente nelle zone attigue.
La velocità della propagazione ---> se la diffusione elettrotonica arriva più lontano, farà scattare lo spyke più lontano. La velocità dello spyke cresce con la larghezza della fibra dell'assone (37%).
Se mielinizzo molte uscite per far andare gli ioni molto più avanti, l'intensità si riduce perché c'è comunque una minima perdita di ioni lungo il tragitto, attraverso la guaina mielinica: quindi la guaina non deve essere troppo lunga e va interrotta sistematicamente ad intervalli regolari, formando tra una guaina e l'altra dei nodi di ranviè ---> grazie a questi, in punti molto piccoli c'è un'intensità molto alta perché concentrata, la soglia viene superata e scatta lo spyke, rigenerando il potenziale d'azione e riaccumolando intensità ---> Conduzione saltatoria
solo gli assoni si possono mielinizzare per via dei canali a voltaggio dipendente
|
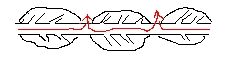
L'ingresso degli ioni di sodio per l'attivazione dei canali è la componente attiva e la diffusione degli ioni quella passiva
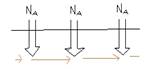 |
La conduzione saltatoria è conveniente anche dal punto di vista metabolico perché avendo una grande forza elettrica in un solo punto, bastano pochi canali per attivare la soglia.
Il potenziale d'azione si scatena per la prima volta nel cono di emergenza (all'attacco dell'assone al soma), con i canali a voltaggio dipendente.
Si innesca una corrente nel dendrite, che elettricamente si diffonde nelle zone circostanti; se essendo decrementale, mantiene un potenziale sufficiente a superare la soglia nel cono, si innesca il potenziale d'azione.
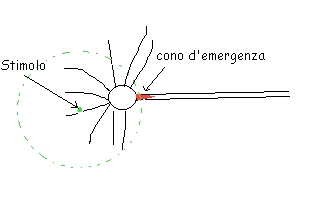 |
Se la corrente è attivata più lontano, è probabile che l'intensità si riduca e la soglia non venga superata: il potenziale d'azione non si innescherà.
Se simultaneamente inserisco corrente in due punti diversi, questi si sommano e, nel caso che da soli non fossero sufficienti ad attivare il potenziale d'azione, ora lo attiveranno. Nel sistema nervoso centrale la sommanza di due correnti è necessaria perché una non basta mai.
Perché la corrente non va all'indietro? Nel soma: perché non ci sono canali a voltaggio dipendente; nell'assone: non viaggia all'indietro perché quando c'è il potenziale d'azione in un punto, in quello precedente c'è appena stato e si è in una fase refrattaria (refrattarietà assoluta --> i canali sono bloccati dalla porzione globulare)
la sclerosi multipla è una malattia demielinizzante ---> senza la mielina c'è la propagazione elettrotonica lentissima (senza canali a voltaggio dipendente), e il potenziale si esaurisce prima di arrivare al nodo di ranviè successivo (con i canali). I primi sintomi sono motori. La coordinazione degli spyke è importantissima, quando è alterata il sistema va in crisi.
Sinapsi ---> parte terminale dell'assone o dei rami dell'assone. È composto da una parte pre-sinaptica (parte terminale dell'assone) e da una parte post-sinaptica (cellula bersaglio), con rispettive membrane pre e post-sinaptiche. Tra la due c'è lo spazio inter-sinaptico.
La sinapsi elettrica ---> il bottone sinaptico, con dei ponti protoplasmatici che collegano i due spazi. Questi ponti sono grosse proteine tuboliformi.
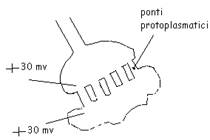 |
In questi punti il potenziale è a + 30 millivolt (altissimo) e innesca sempre uno spyke talmente forte da mandare il segnale oltre la soglia nel cono d'emergenza (spyke post-sinaptico), nonostante nei dendriti e nel soma non ci siano canali a voltaggio dipendente.
Questo tipo di sinapsi è diffuso nel regno animale, per via della sua velocità di propagazione; è capace di trasmettere funzioni stereotipate (non modulabili ---> es. tocchi la coda al pesce e scappa immediatamente); è presente per solo il 10% circa del nostro sistema nervoso centrale.
Con questo tipo di sinapsi si è soggetti a risposte sincrone: il segnale viene trasmesso da un neurone a tutti quelli a cui è collegato, nello stesso momento (amplificazione del segnale).
 |
È diffuso nel regno degli invertebrati.
La sinapsi chimica ---> è sostanzialmente l'opposto. Nell'assone viaggiano le vescicole sinaptiche, al cui interno sono presenti i neurotrasmettitori. Nella sinapsi, oltre ai canali a voltaggio dipendente per il sodio, si aprono al raggiungimento della soglia di potenziale anche i canali per il calcio, che entra nella membrana pre-sinaptica. Questo, non è ben chiaro come, attira le vescicole verso la membrana, e al contatto con essa si fonderanno (rilasciando i neurotrasmettitori nella sinapsi), per poi essere riassorbite.
Molecola ---> combinazione di atomi
Proteina ---> combinazione di decine di migliaia di atomi
Amminoacido ---> combinazione di decine-centinaia di migliaia di atomi
Il neurotrasmettitore è un amminoacido
I neurotrasmettitori vanno ad agire sulla membrana post-sinaptica e stimolano i recettori, che, al contatto con questi, si aprono e fanno passare il sodio (in entrata) e/o il potassio (in uscita).
Il sodio entra nella membrana post-sinaptica e fa una depolarizzazione.
Recettori ---> proteine di membrana, può essere un canale diretto o no: quando non è diretto, ci può essere bisogno di una serie di reazioni chimiche per farlo aprire e far passare sodio e/o potassio.
Neurotrasmissione ---> quando i recettori e canale sono la stessa cosa
Neurotrasmissione modulare ---> quando sono separati
|
Nei collegamenti con le cellule muscolari, il sodio che entra inizialmente nelle membrana post-sinaptica, porta il potenziale a valori più alti, attivando i canali a voltaggio dipendente per il sodio, che a loro volta faranno entrare più sodio e aumentare il potenziale, in un ciclo autorigenerante fino a che si crea un picco d'azione tale da far contrarre il muscolo.
Di solito i canali sono selettivi per il sodio, altre volte per il potassio ecc... A seconda del tipo di specie ionica facilitata nell'ingresso, ci saranno effetti diversi.
Nella membrana post-sinaptica ci sono migliaia di canali per il sodio, una volta entrate queste depolarizzano la membrana (Potenziale Post-Sinaptico Eccitatorio [PPSE]), aumentando il potenziale. Eccitatorio perché se il segnale è capace di raggiungere il cono d'emergenza, c'è la probabilità di attivazione dello spyke.
Se il neurotrasmettitore ha l'effetto di chiudere i canali per il sodio, provoca una iperpolarizzazione (il potenziale diminuisce: -70, -80...) e si chiama Potenziale Post-Sinaptico Inibitorio (PPSI).
Il fatto che sia eccitatorio o inibitorio dipende dall'effetto finale che si ha sul canale:
effetto finale depolarizzante ---> eccitatorio
effetto finale iperpolarizzante ---> inibitorio
Non è possibile avere tutti e due gli effetti contemporaneamente nelle varie sinapsi dello stesso neurone: ci saranno quindi neuroni o eccitatori o inibitori.
Sommazione spaziale ---> quando si sommano due segnali provenienti due dendriti (quindi distanti nello spazio)
Sommazione temporale ---> quando arrivano due spyke dalla stessa sinapsi (prima uno e poi l'altro), ma devono essere sufficientemente ravvicinati nel tempo.
Fonte: http://appunti.buzzionline.eu/downloads/neurobiologia0506.doc
Autori: Sara e Stefano
Neurobiologia cellulare
Visita la nostra pagina principale
Neurobiologia cellulare
Termini d' uso e privacy